

DNA helicase-mediated packaging of adeno-associated virus type 2 genomes into preformed capsids
- PMID: 11406604
- PMCID: PMC150213
- DOI: 10.1093/emboj/20.12.3282
DNA helicase-mediated packaging of adeno-associated virus type 2 genomes into preformed capsids
Abstract
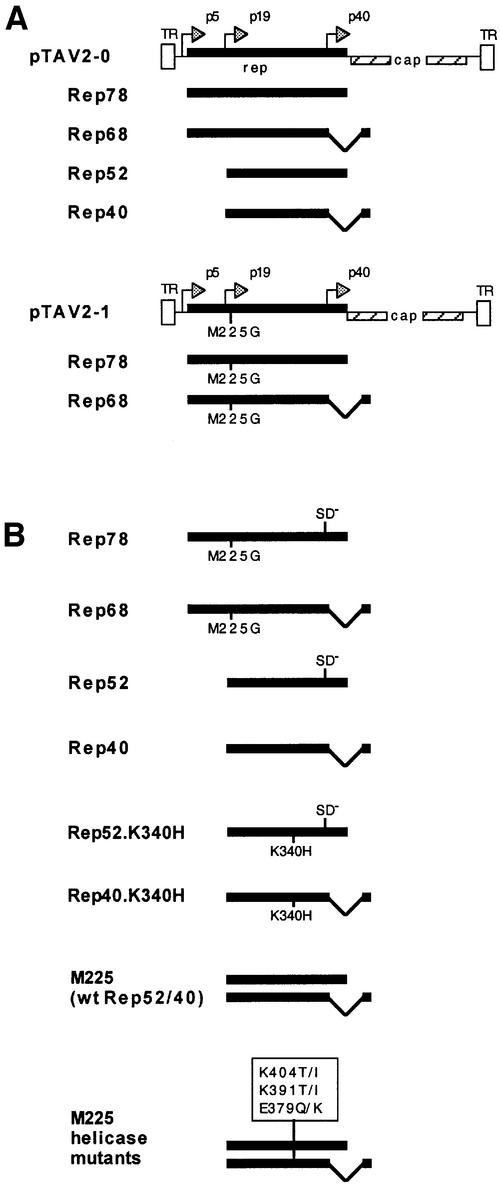
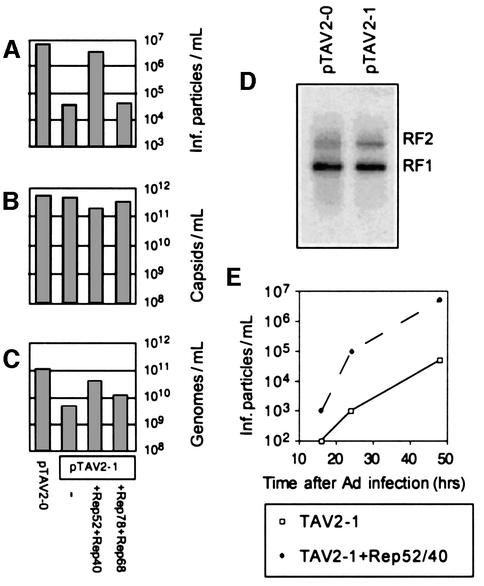
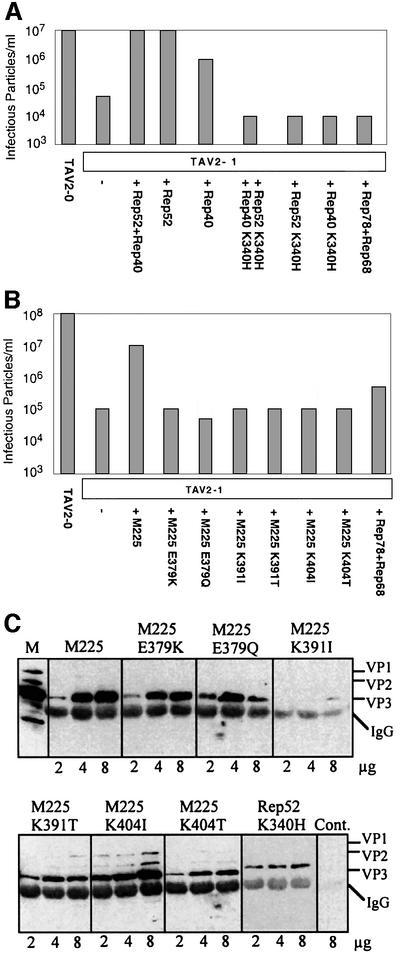
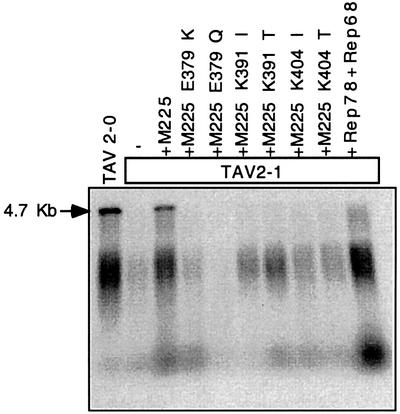
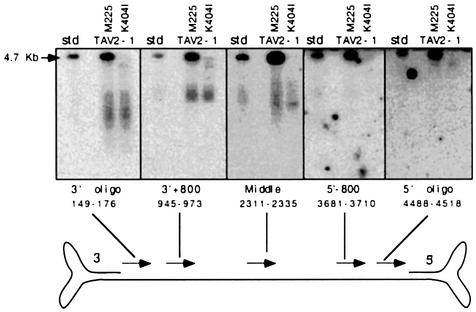
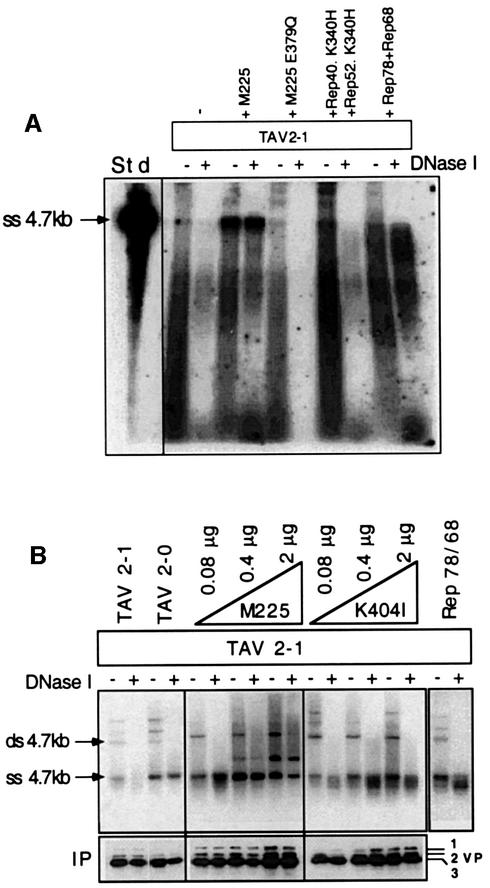
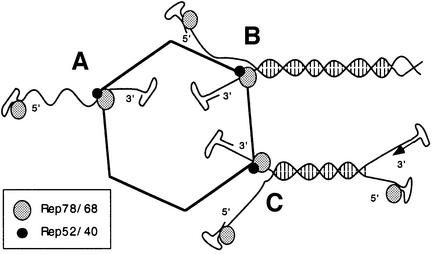
Helicases not only catalyse the disruption of hydrogen bonding between complementary regions of nucleic acids, but also move along nucleic acid strands in a polar fashion. Here we show that the Rep52 and Rep40 proteins of adeno-associated virus type 2 (AAV-2) are required to translocate capsid-associated, single-stranded DNA genomes into preformed empty AAV-2 capsids, and that the DNA helicase function of Rep52/40 is essential for this process. Furthermore, DNase protection experiments suggest that insertion of AAV-2 genomes proceeds from the 3' end, which correlates with the 3'-->5' processivity demonstrated for the Rep52/40 helicase. A model is proposed in which capsid-immobilized helicase complexes act as molecular motors to 'pump' single-stranded DNA across the capsid boundary.
Figures
References
-
- Baker T.A. and Bell,S.P. (1998) Polymerases and the replisome: machines within machines. Cell, 92, 295–305. - PubMed
-
- Bina M., Ng,S.C. and Blasquez,V. (1983) Simian virus 40 chromatin interaction with the capsid proteins. J. Biomol. Struct. Dyn., 1, 689–704. - PubMed
-
- Bird L.E., Subramanya,H.S. and Wigley,D.B. (1998) Helicases: a unifying structural theme? Curr. Opin. Struct. Biol., 8, 14–18. - PubMed
-
- Black L.W. (1989) DNA packaging in dsDNA bacteriophages. Annu. Rev. Microbiol., 43, 267–292. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
