

The polo-like kinase Plx1 is required for activation of the phosphatase Cdc25C and cyclin B-Cdc2 in Xenopus oocytes
- PMID: 11408585
- PMCID: PMC37341
- DOI: 10.1091/mbc.12.6.1791
The polo-like kinase Plx1 is required for activation of the phosphatase Cdc25C and cyclin B-Cdc2 in Xenopus oocytes
Abstract
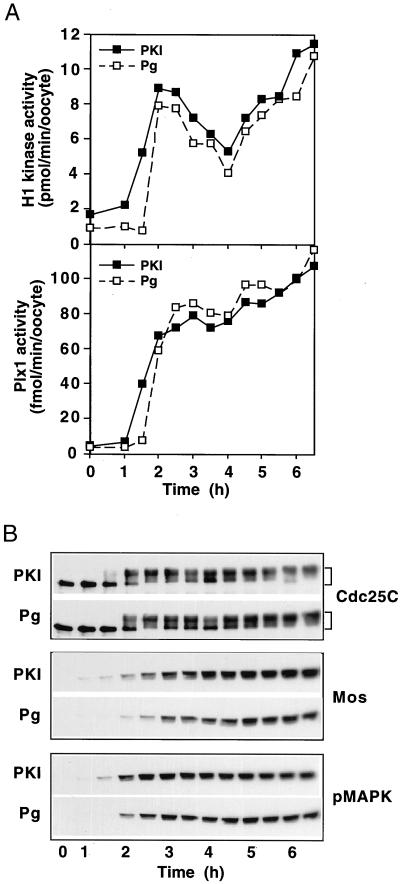
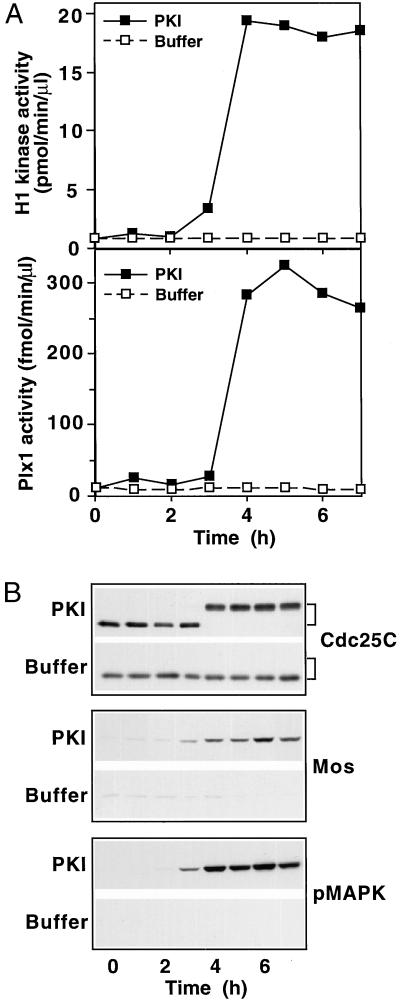
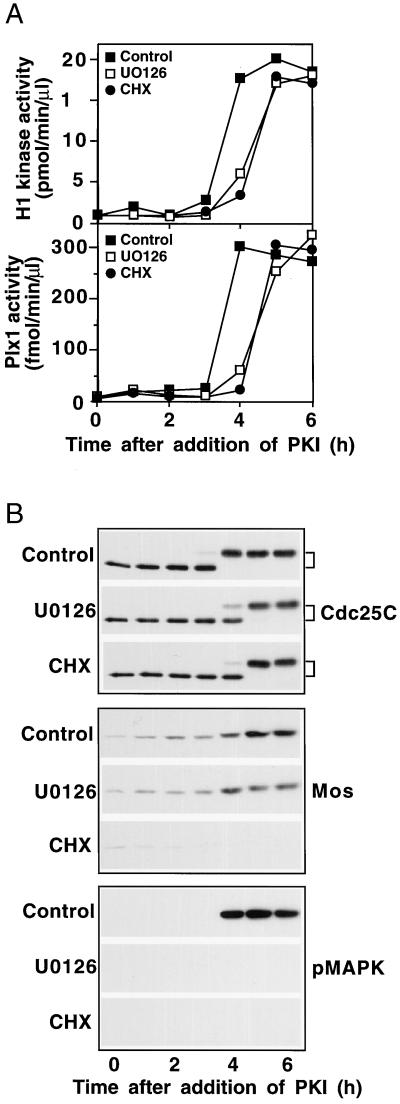
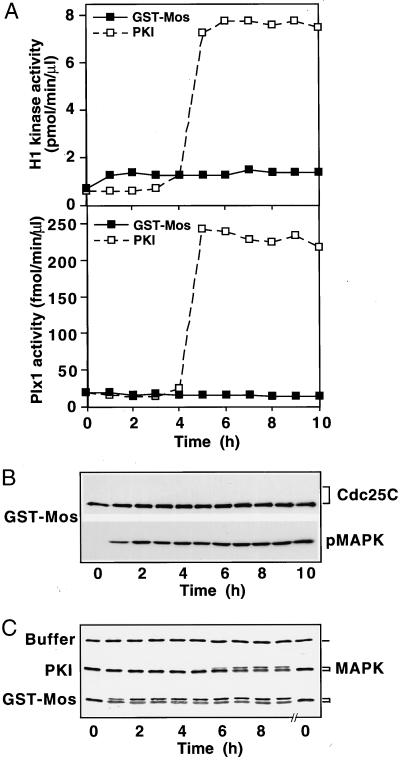
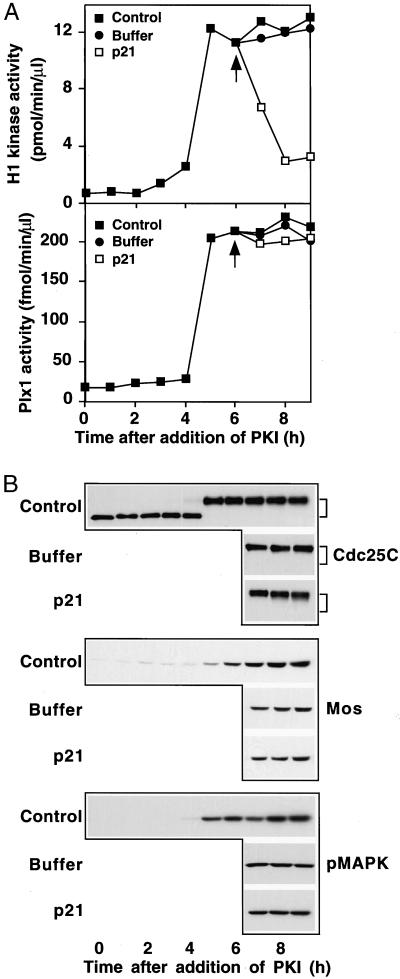
In the Xenopus oocyte system mitogen treatment triggers the G(2)/M transition by transiently inhibiting the cAMP-dependent protein kinase (PKA); subsequently, other signal transduction pathways are activated, including the mitogen-activated protein kinase (MAPK) and polo-like kinase pathways. To study the interactions between these pathways, we have utilized a cell-free oocyte extract that carries out the signaling events of oocyte maturation after addition of the heat-stable inhibitor of PKA, PKI. PKI stimulated the synthesis of Mos and activation of both the MAPK pathway and the Plx1/Cdc25C/cyclin B-Cdc2 pathway. Activation of the MAPK pathway alone by glutathione S-transferase (GST)-Mos did not lead to activation of Plx1 or cyclin B-Cdc2. Inhibition of the MAPK pathway in the extract by the MEK1 inhibitor U0126 delayed, but did not prevent, activation of the Plx1 pathway, and inhibition of Mos synthesis by cycloheximide had a similar effect, suggesting that MAPK activation is the only relevant function of Mos. Immunodepletion of Plx1 completely inhibited activation of Cdc25C and cyclin B-Cdc2 by PKI, indicating that Plx1 is necessary for Cdc25C activation. In extracts containing fully activated Plx1 and Cdc25C, inhibition of cyclin B-Cdc2 by p21(Cip1) had no significant effect on either the phosphorylation of Cdc25C or the activity of Plx1. These results demonstrate that maintenance of Plx1 and Cdc25C activity during mitosis does not require cyclin B-Cdc2 activity.
Figures

Similar articles
-
Activated polo-like kinase Plx1 is required at multiple points during mitosis in Xenopus laevis.Mol Cell Biol. 1998 Jul;18(7):4262-71. doi: 10.1128/MCB.18.7.4262. Mol Cell Biol. 1998. PMID: 9632810 Free PMC article.
-
The Polo-like kinase Plx1 is a component of the MPF amplification loop at the G2/M-phase transition of the cell cycle in Xenopus eggs.J Cell Sci. 1998 Jun;111 ( Pt 12):1751-7. doi: 10.1242/jcs.111.12.1751. J Cell Sci. 1998. PMID: 9601104
-
Cdc2-cyclin B triggers H3 kinase activation of Aurora-A in Xenopus oocytes.J Biol Chem. 2003 Jun 13;278(24):21439-49. doi: 10.1074/jbc.M300811200. Epub 2003 Apr 1. J Biol Chem. 2003. PMID: 12670933
-
New pathways from PKA to the Cdc2/cyclin B complex in oocytes: Wee1B as a potential PKA substrate.Cell Cycle. 2006 Feb;5(3):227-31. doi: 10.4161/cc.5.3.2395. Epub 2006 Feb 8. Cell Cycle. 2006. PMID: 16418576 Review.
-
Xenopus Polo-like kinase Plx1: a multifunctional mitotic kinase.Oncogene. 2005 Jan 10;24(2):238-47. doi: 10.1038/sj.onc.1208220. Oncogene. 2005. PMID: 15640839 Review.
Cited by
-
Cdk1 activity is required for mitotic activation of aurora A during G2/M transition of human cells.J Biol Chem. 2010 Jul 9;285(28):21849-57. doi: 10.1074/jbc.M110.141010. Epub 2010 May 5. J Biol Chem. 2010. PMID: 20444701 Free PMC article.
-
Targeting Echinococcus multilocularis stem cells by inhibition of the Polo-like kinase EmPlk1.PLoS Negl Trop Dis. 2014 Jun 5;8(6):e2870. doi: 10.1371/journal.pntd.0002870. eCollection 2014 Jun. PLoS Negl Trop Dis. 2014. PMID: 24901228 Free PMC article.
-
Pioneering the Xenopus oocyte and egg extract system.J Biol Chem. 2012 Jun 22;287(26):21640-53. doi: 10.1074/jbc.X112.371161. Epub 2012 May 8. J Biol Chem. 2012. PMID: 22570479 Free PMC article. No abstract available.
-
Normal cell cycle and checkpoint responses in mice and cells lacking Cdc25B and Cdc25C protein phosphatases.Mol Cell Biol. 2005 Apr;25(7):2853-60. doi: 10.1128/MCB.25.7.2853-2860.2005. Mol Cell Biol. 2005. PMID: 15767688 Free PMC article.
-
PP1 control of M phase entry exerted through 14-3-3-regulated Cdc25 dephosphorylation.EMBO J. 2003 Nov 3;22(21):5734-45. doi: 10.1093/emboj/cdg545. EMBO J. 2003. PMID: 14592972 Free PMC article.
References
-
- Dunphy WG, Kumagai A. The cdc25 protein contains an intrinsic phosphatase activity. Cell. 1991;67:189–196. - PubMed
-
- Favata F, Horiuchi YY, Manos EJ, Daulerio AJ, Stradley DA, Feeser WS, Van Dyk DE, Pitts WJ, Earl RA, Hobbs F, Copeland RA, Magolda RL, Scherle PA, Trzaskos JM. Identification of a novel inhibitor of mitogen-activated protein kinase kinase. J Biol Chem. 1998;273:18623–18632. - PubMed
-
- Fisher DL, Brassac T, Galas S, Doree M. Dissociation of MAP kinase activation and MPF activation in hormone-stimulated maturation of Xenopus oocytes. Development. 1999;126:4537–4546. - PubMed
-
- Gautier J, Solomon MJ, Booher RN, Bazan JF, Kirschner MW. cdc25 is a specific tyrosine phosphatase that directly activates p34cdc2. Cell. 1991;67:197–211. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
