

Regulation of epidermal growth factor receptor signaling by endocytosis and intracellular trafficking
- PMID: 11408594
- PMCID: PMC37350
- DOI: 10.1091/mbc.12.6.1897
Regulation of epidermal growth factor receptor signaling by endocytosis and intracellular trafficking
Abstract
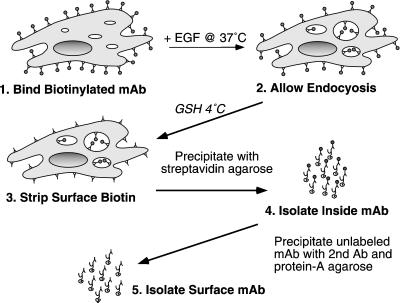
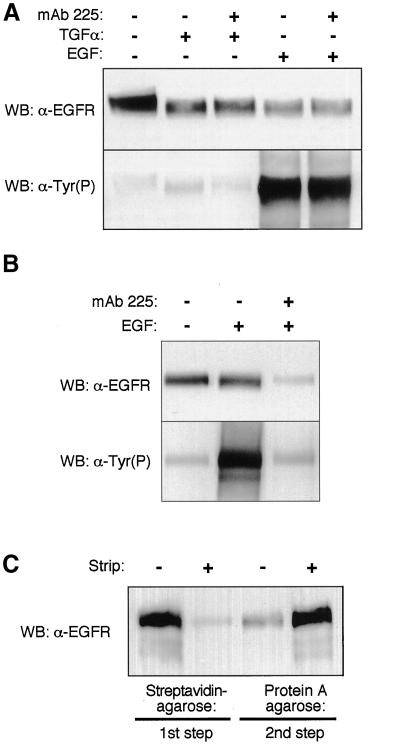
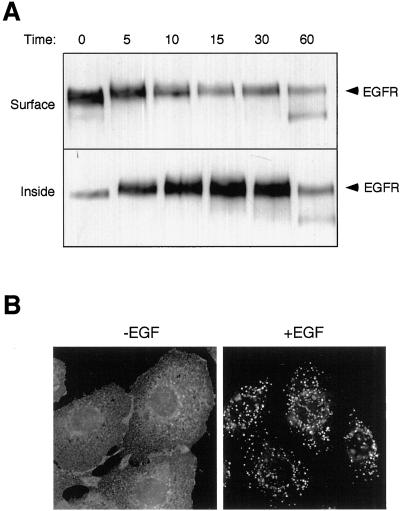
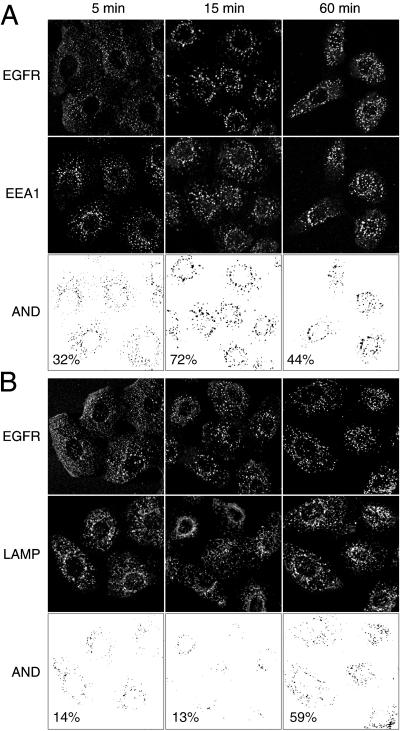
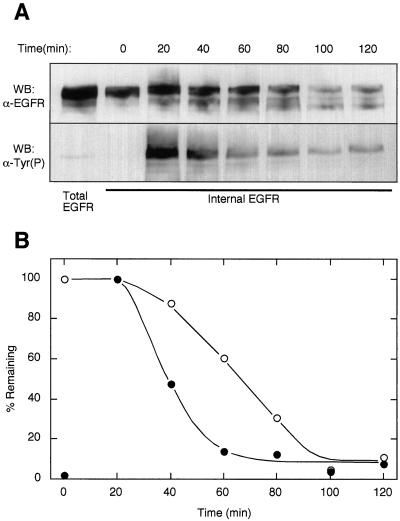
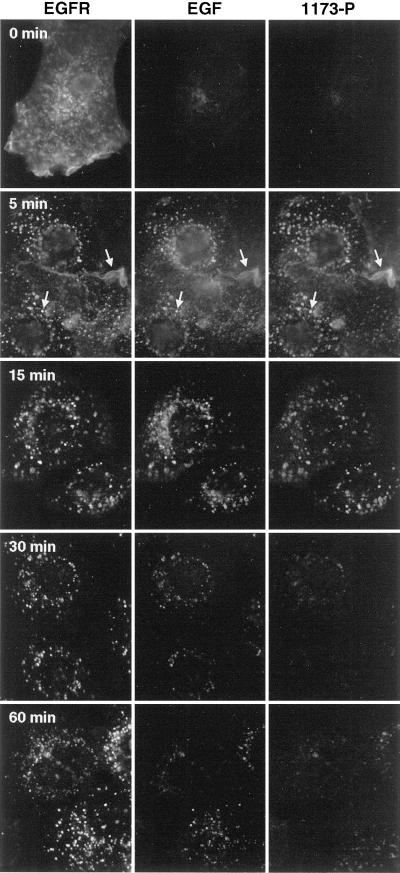
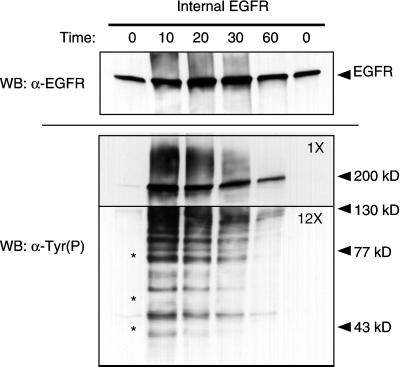
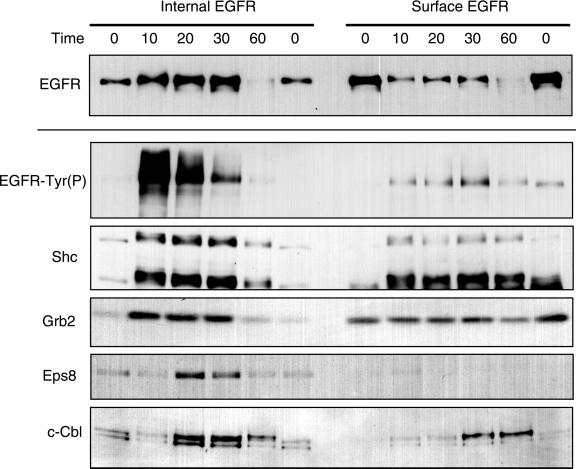
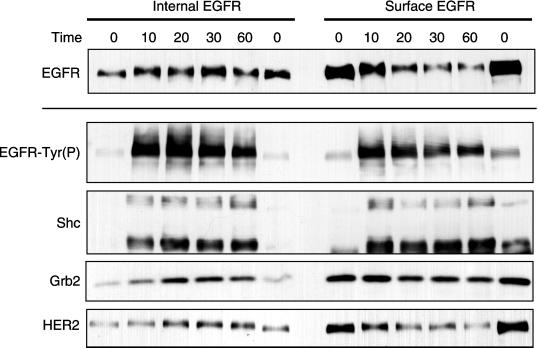
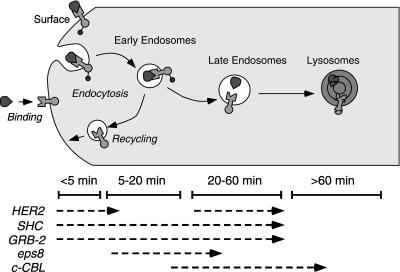
Ligand activation of the epidermal growth factor receptor (EGFR) leads to its rapid internalization and eventual delivery to lysosomes. This process is thought to be a mechanism to attenuate signaling, but signals could potentially be generated after endocytosis. To directly evaluate EGFR signaling during receptor trafficking, we developed a technique to rapidly and selectively isolate internalized EGFR and associated molecules with the use of reversibly biotinylated anti-EGFR antibodies. In addition, we developed antibodies specific to tyrosine-phosphorylated EGFR. With the use of a combination of fluorescence imaging and affinity precipitation approaches, we evaluated the state of EGFR activation and substrate association during trafficking in epithelial cells. We found that after internalization, EGFR remained active in the early endosomes. However, receptors were inactivated before degradation, apparently due to ligand removal from endosomes. Adapter molecules, such as Shc, were associated with EGFR both at the cell surface and within endosomes. Some molecules, such as Grb2, were primarily found associated with surface EGFR, whereas others, such as Eps8, were found only with intracellular receptors. During the inactivation phase, c-Cbl became EGFR associated, consistent with its postulated role in receptor attenuation. We conclude that the association of the EGFR with different proteins is compartment specific. In addition, ligand loss is the proximal cause of EGFR inactivation. Thus, regulated trafficking could potentially influence the pattern as well as the duration of signal transduction.
Figures
References
-
- Ahn S, Maudsley S, Luttrell LM, Lefkowitz RJ, Daaka Y. Src-mediated tyrosine phosphorylation of dynamin is required for beta2- adrenergic receptor internalization and mitogen-activated protein kinase signaling. J Biol Chem. 1999;274:1185–1188. - PubMed
-
- Aroian RV, Koga M, Mendel JE, Ohshima Y, Sternberg PW. The let-23 gene necessary for Caenorhabditis elegans vulval induction encodes a tyrosine kinase of the EGF receptor subfamily. Nature. 1990;348:693–699. - PubMed
-
- Baass PC, Guglielmo FA, Posner BI, Bergeron JM. Compartmentalized signal transduction by receptor tyrosine kinases. Trends Cell Biol. 1995;5:465–470. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
