

RNA recombination between persisting pestivirus and a vaccine strain: generation of cytopathogenic virus and induction of lethal disease
- PMID: 11413291
- PMCID: PMC114347
- DOI: 10.1128/JVI.75.14.6256-6264.2001
RNA recombination between persisting pestivirus and a vaccine strain: generation of cytopathogenic virus and induction of lethal disease
Abstract
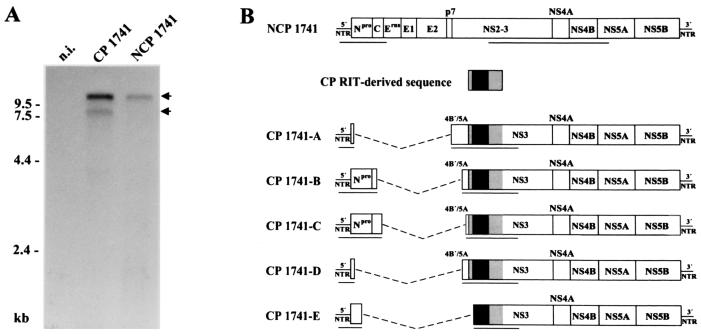
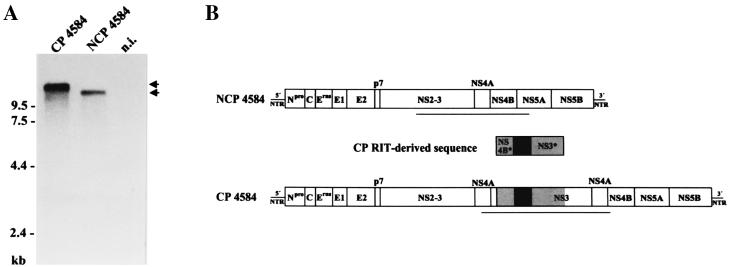
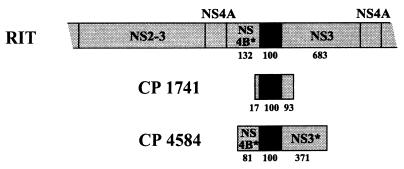
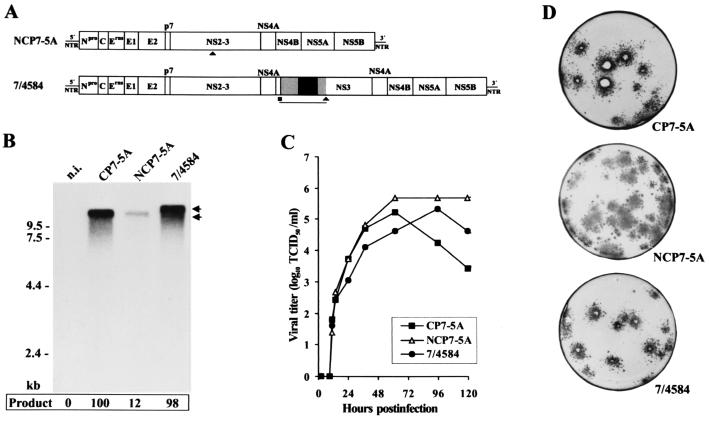
Molecular analysis of a cytopathogenic (cp) bovine viral diarrhea virus (BVDV) isolate (1741) obtained from a case of mucosal disease (MD) led to the identification of five different viral subgenomic RNAs in addition to a noncytopathogenic (noncp) strain (NCP 1741). For each of the subgenomes, a large internal deletion was found together with an inserted sequence encoding part of ribosomal protein S27a fused to an N-terminally truncated ubiquitin monomer. Surprisingly, the two cellular insertions together with flanking viral sequences encoding parts of NS3 and NS4B are >99% identical to the previously described sequence of BVDV vaccine strain RIT (P. Becher, M. Orlich, and H.-J. Thiel, J. Virol. 72:8697-8704, 1998), while the remainder of the subgenomes is derived from the genome of NCP 1741. Further analyses including molecular cloning and nucleotide sequencing of the recombination partners revealed that both homologous and nonhomologous RNA recombination contributed to the generation of the viral subgenomes. Interestingly, for another cp BVDV isolate (CP 4584) from an independent case of MD, again an insertion of a RIT-derived sequence element was detected. In contrast to CP 1741, for CP 4584 a duplication of the genomic region encoding NS3 and parts of NS4A and NS4B was found. Transfection of bovine cells with RNA transcribed from a chimeric cDNA construct showed that the RIT-derived insertion together with the CP 4584-specific duplication of viral sequences represents the genetic basis of cytopathogenicity of CP 4584. Remarkably, passages of the recovered cp virus in cell culture led to emergence of noncp BVDV and a number of viral subgenomes whose genome organization was similar to that in BVDV 1741.
Figures
References
-
- Baker J C. Bovine viral diarrhea virus: a review. J Am Vet Med Assoc. 1987;190:1449–1458. - PubMed
-
- Baroth M, Orlich M, Thiel H-J, Becher P. Insertion of cellular NEDD8 coding sequences in a pestivirus. Virology. 2000;278:456–466. - PubMed
-
- Becher P, Orlich M, Shannon A D, Horner G, König M, Thiel H-J. Phylogenetic analysis of pestiviruses from domestic and wild ruminants. J Gen Virol. 1997;78:1357–1366. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
