

Gamma -secretase inhibitors repress thymocyte development
- PMID: 11416218
- PMCID: PMC34695
- DOI: 10.1073/pnas.131202798
Gamma -secretase inhibitors repress thymocyte development
Abstract
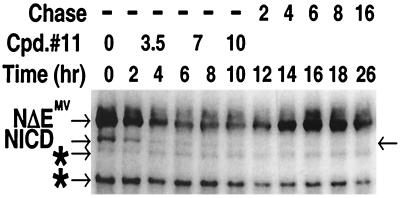
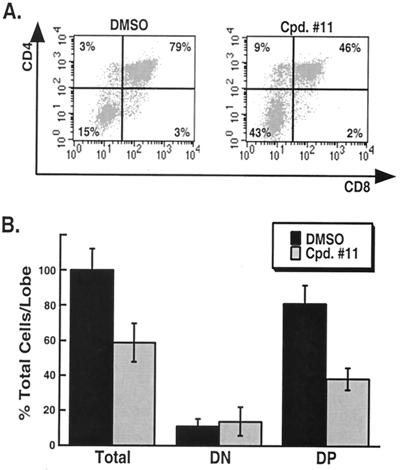
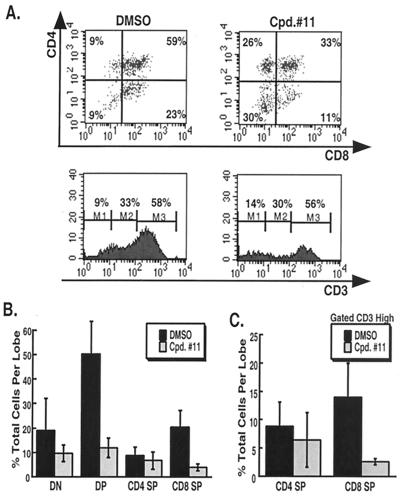
A major therapeutic target in the search for a cure to the devastating Alzheimer's disease is gamma-secretase. This activity resides in a multiprotein enzyme complex responsible for the generation of Abeta42 peptides, precipitates of which are thought to cause the disease. Gamma-secretase is also a critical component of the Notch signal transduction pathway; Notch signals regulate development and differentiation of adult self-renewing cells. This has led to the hypothesis that therapeutic inhibition of gamma-secretase may interfere with Notch-related processes in adults, most alarmingly in hematopoiesis. Here, we show that application of gamma-secretase inhibitors to fetal thymus organ cultures interferes with T cell development in a manner consistent with loss or reduction of Notch1 function. Progression from an immature CD4-/CD8- state to an intermediate CD4+/CD8+ double-positive state was repressed. Furthermore, treatment beginning later at the double-positive stage specifically inhibited CD8+ single-positive maturation but did not affect CD4+ single-positive cells. These results demonstrate that pharmacological gamma-secretase inhibition recapitulates Notch1 loss in a vertebrate tissue and present a system in which rapid evaluation of gamma-secretase-targeted pharmaceuticals for their ability to inhibit Notch activity can be performed in a relevant context.
Figures
References
-
- Artavanis-Tsakonas S, Rand M D, Lake R J. Science. 1999;284:770–776. - PubMed
-
- Milner L A, Bigas A. Blood. 1999;93:2431–2448. - PubMed
-
- Deftos M L, Bevan M J. Curr Opin Immunol. 2000;12:166–172. - PubMed
-
- Mumm J S, Schroeter E H, Saxena M T, Griesemer A, Tian X, Pan D J, Ray W J, Kopan R. Mol Cell. 2000;5:197–206. - PubMed
-
- Struhl G, Greenwald I. Nature (London) 1999;398:522–525. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
