

Identification and characterization of a melanin-concentrating hormone receptor
- PMID: 11416225
- PMCID: PMC34710
- DOI: 10.1073/pnas.131200698
Identification and characterization of a melanin-concentrating hormone receptor
Abstract
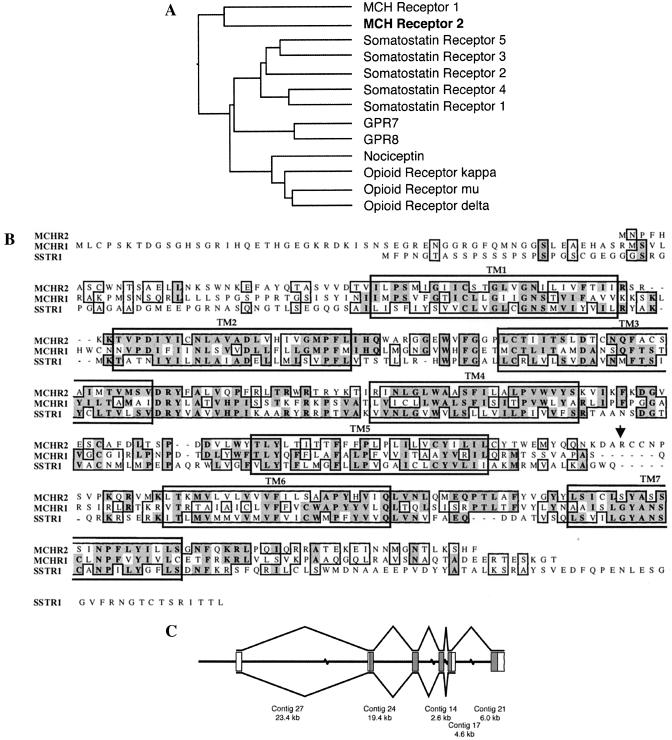
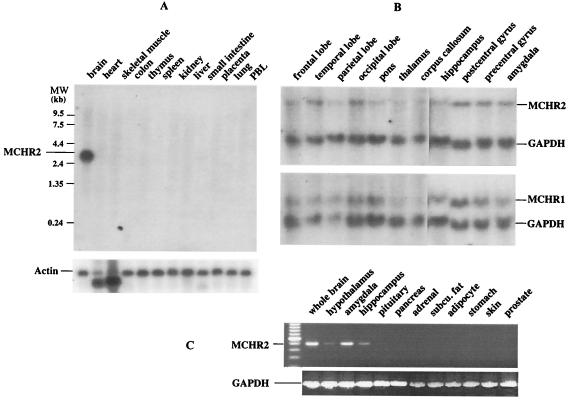
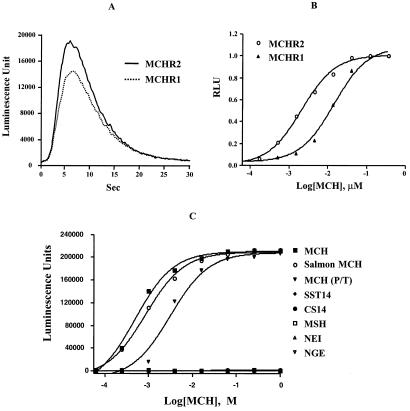
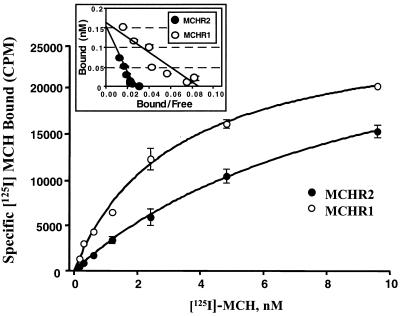
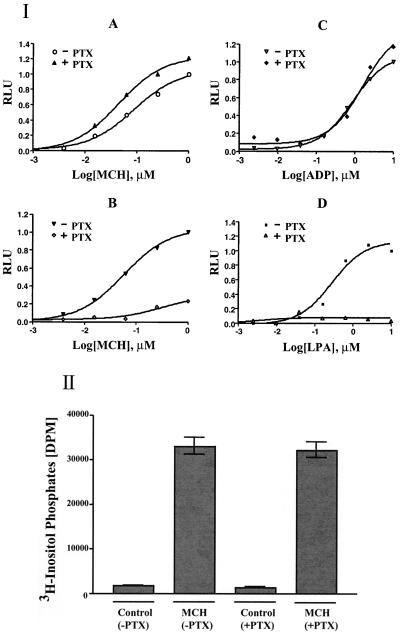
Melanin-concentrating hormone (MCH), a neuropeptide expressed in central and peripheral nervous systems, plays an important role in the control of feeding behaviors and energy metabolism. An orphan G protein-coupled receptor (SLC-1/GPR24) has recently been identified as a receptor for MCH (MCHR1). We report here the identification and characterization of a G protein-coupled receptor as the MCH receptor subtype 2 (MCHR2). MCHR2 has higher protein sequence homology to MCHR1 than any other G protein-coupled receptor. The expression of MCHR2 has been detected in many regions of the brain. In contrast to MCHR1, which is intronless in the coding region and is located at the chromosomal locus 22q13.3, the MCHR2 gene has multiple exons and is mapped to locus 6q21. MCHR2 is specifically activated by nanomolar concentrations of MCH, binds to MCH with high affinity, and signals through Gq protein. This discovery is important for a full understanding of MCH biology and the development of potential therapeutics for diseases involving MCH, including obesity.
Figures
References
-
- Kawauchi H, Kawazoe I, Tsubokawa M, Kishida M, Baker B I. Nature (London) 1983;305:321–323. - PubMed
-
- Skofitsch G, Jacobowitz D M, Zamir N. Brain Res Bull. 1985;15:635–649. - PubMed
-
- Bittencourt J C, Presse F, Arias C, Peto C, Vaughan J, Nahon J L, Vale W, Sawchenko P E. J Comp Neurol. 1992;319:218–245. - PubMed
-
- Saito Y, Nothacker H-P, Civelli O. Trends Endocrinol Metabol. 2000;11:299–303. - PubMed
-
- Qu D, Ludwig D S, Gammeltoft S, Piper M, Pelleymounter M A, Cullen M J, Mathes W G, Przypek J, Kanarek R, Maratos-Flier E. Nature (London) 1996;380:243–247. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
