

Constitutive septal murein synthesis in Escherichia coli with impaired activity of the morphogenetic proteins RodA and penicillin-binding protein 2
- PMID: 11418550
- PMCID: PMC95299
- DOI: 10.1128/JB.183.14.4115-4126.2001
Constitutive septal murein synthesis in Escherichia coli with impaired activity of the morphogenetic proteins RodA and penicillin-binding protein 2
Abstract
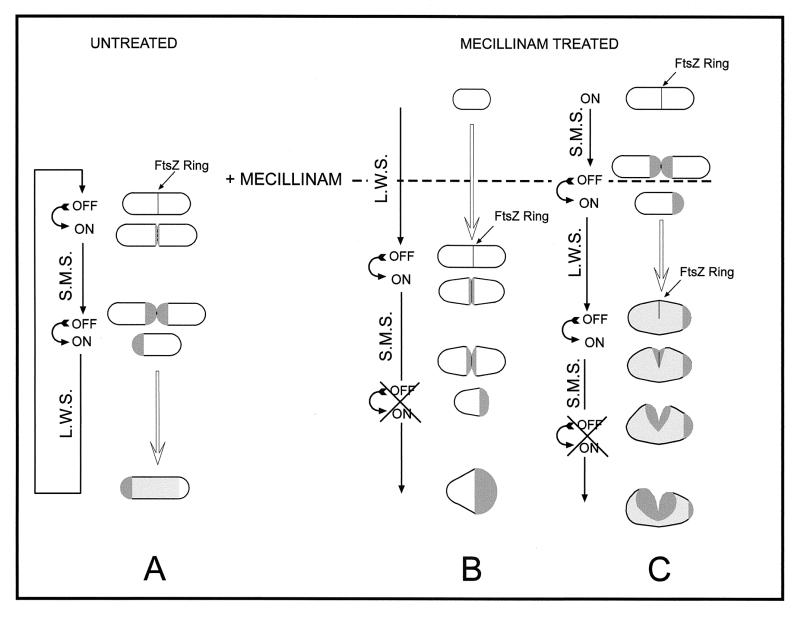
The pattern of peptidoglycan (murein) segregation in cells of Escherichia coli with impaired activity of the morphogenetic proteins penicillin-binding protein 2 and RodA has been investigated by the D-cysteine-biotin immunolabeling technique (M. A. de Pedro, J. C. Quintela, J.-V. Höltje, and H. Schwarz, J. Bacteriol. 179:2823-2834, 1997). Inactivation of these proteins either by amdinocillin treatment or by mutations in the corresponding genes, pbpA and rodA, respectively, leads to the generation of round, osmotically stable cells. In normal rod-shaped cells, new murein precursors are incorporated all over the lateral wall in a diffuse manner, being mixed up homogeneously with preexisting material, except during septation, when strictly localized murein synthesis occurs. In contrast, in rounded cells, incorporation of new precursors is apparently a zonal process, localized at positions at which division had previously taken place. Consequently, there is no mixing of new and old murein. Old murein is preserved for long periods of time in large, well-defined areas. We propose that the observed patterns are the result of a failure to switch off septal murein synthesis at the end of septation events. Furthermore, the segregation results confirm that round cells of rodA mutants do divide in alternate, perpendicular planes as previously proposed (K. J. Begg and W. D. Donachie, J. Bacteriol. 180:2564-2567, 1998).
Figures
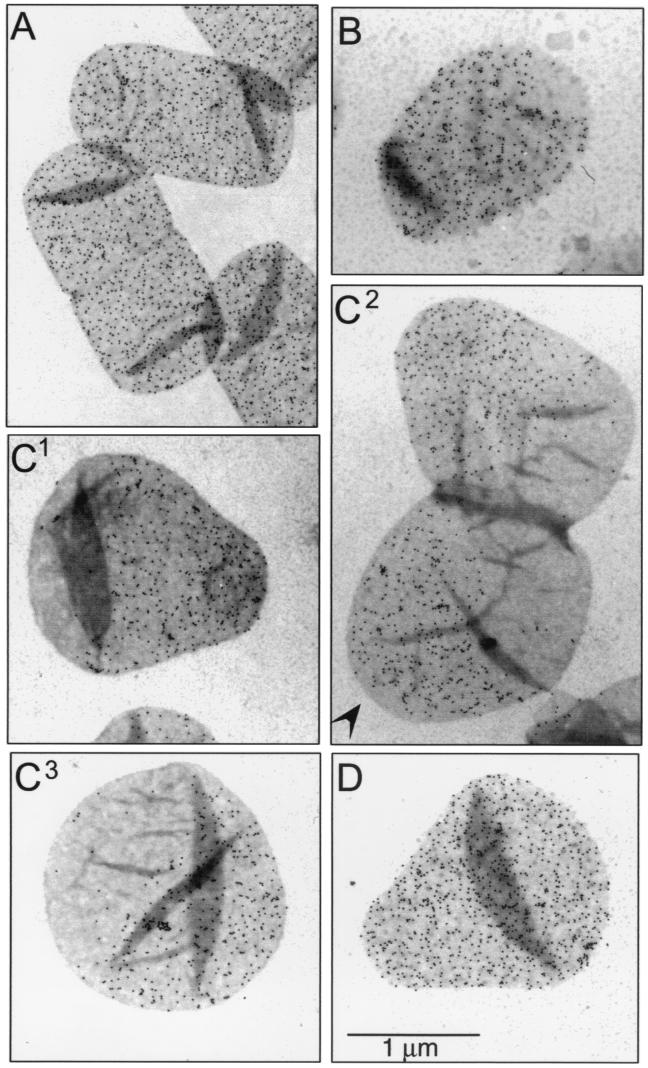
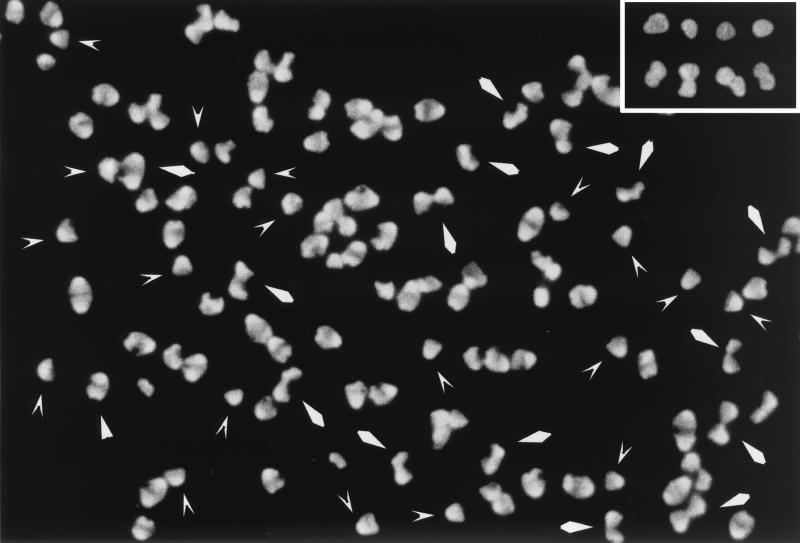
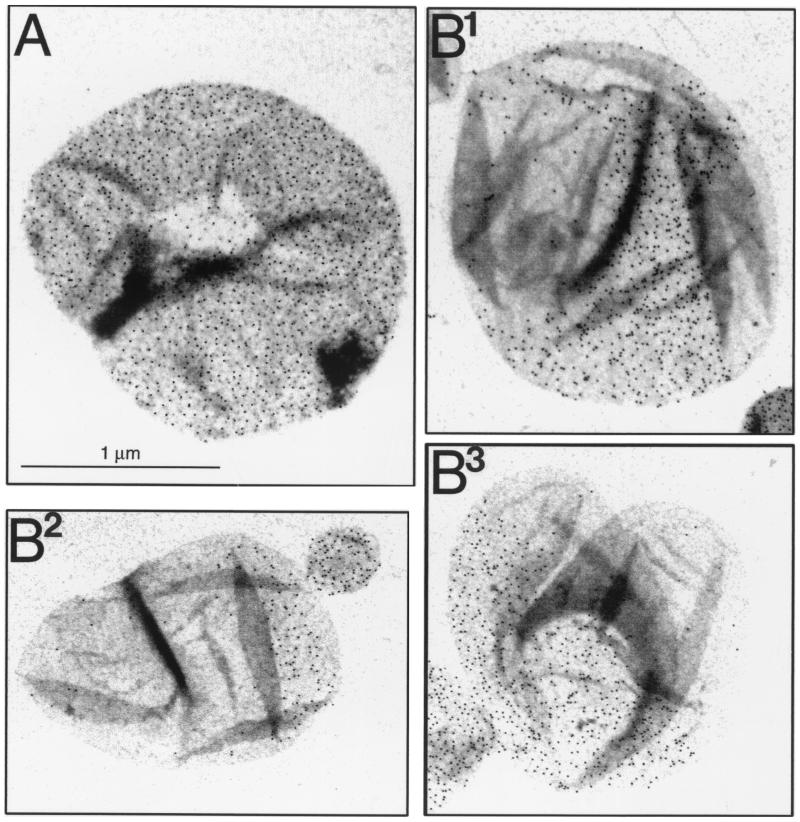

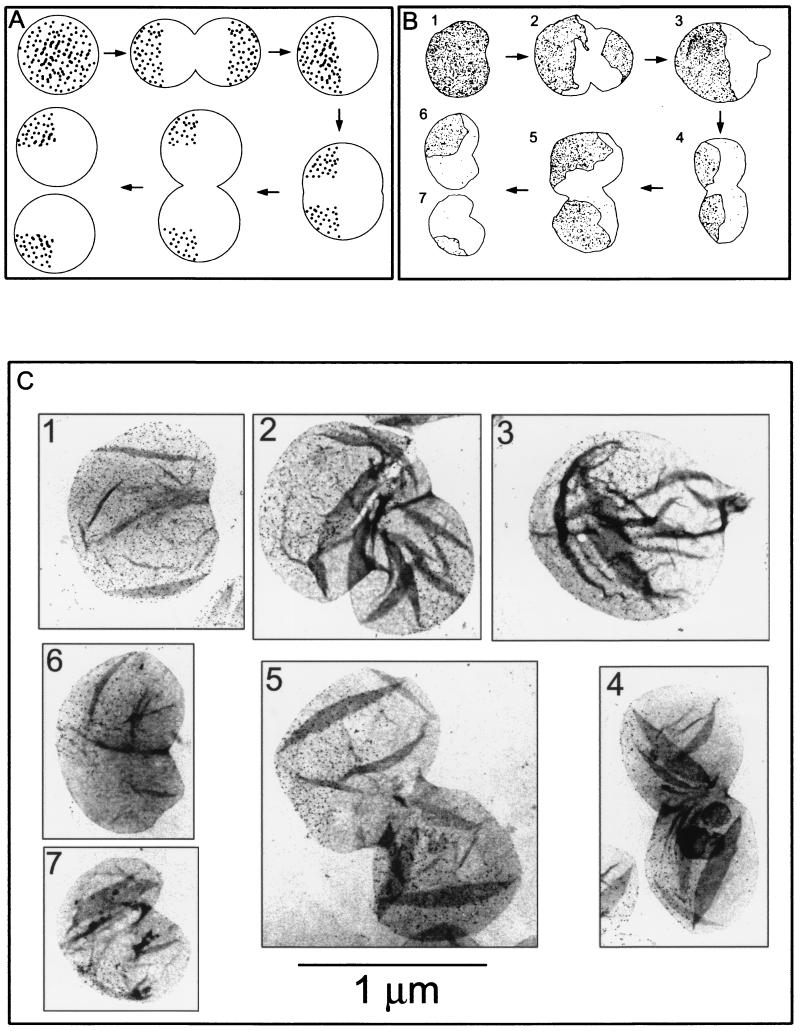
Comment in
-
New insights into the developmental history of the bacterial cell division site.J Bacteriol. 2003 Feb;185(4):1125-7. doi: 10.1128/JB.185.4.1125-1127.2003. J Bacteriol. 2003. PMID: 12562780 Free PMC article. No abstract available.
References
-
- Addinall S G, Lutkenhaus J. FtsZ-spirals and -arcs determine the shape of the invaginating septa in some mutants of Escherichia coli. Mol Microbiol. 1996;22:231–237. - PubMed
-
- Ayala J A, Garrido T, de Pedro M A, Vicente M. Molecular biology of bacterial septation. In: Ghuysen J M, Hakenbeck R, editors. Bacterial cell wall. Amsterdam, The Netherlands: Elsevier; 1994. pp. 73–101.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
