

A critical investigation of the Tanford-Kirkwood scheme by means of Monte Carlo simulations
- PMID: 11420443
- PMCID: PMC2374116
- DOI: 10.1110/ps.42601
A critical investigation of the Tanford-Kirkwood scheme by means of Monte Carlo simulations
Abstract
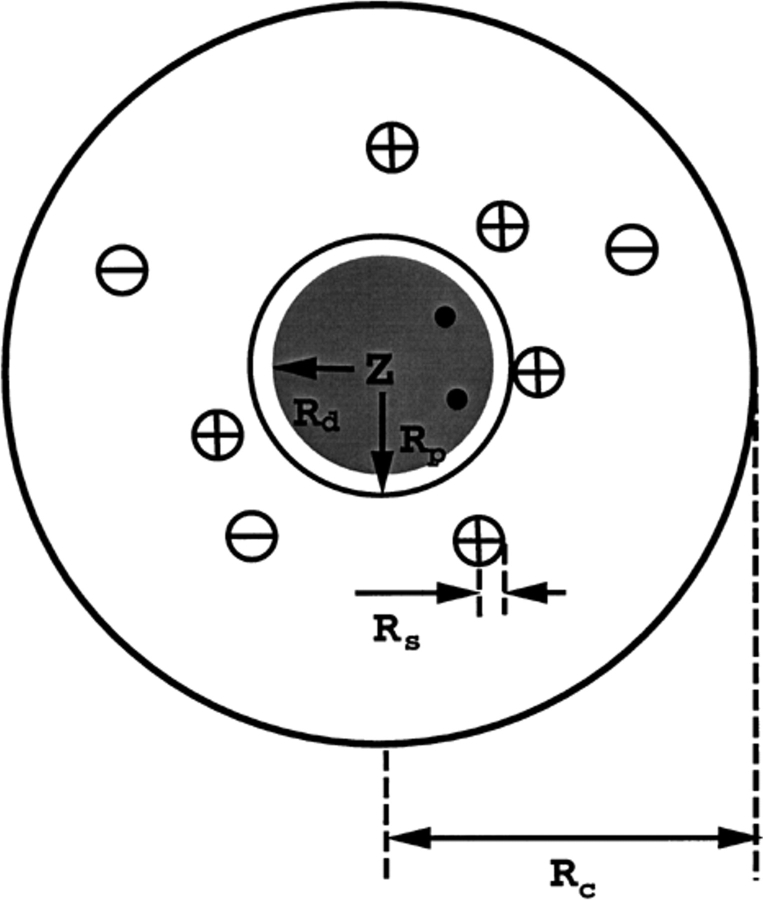
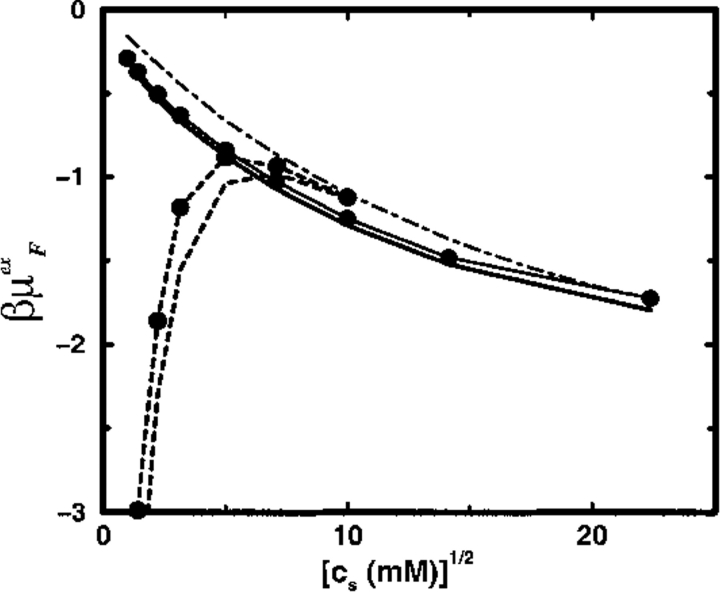
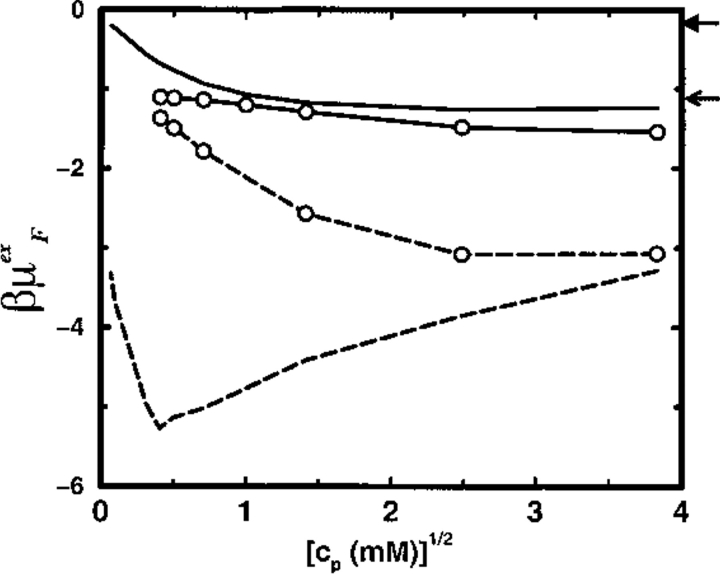
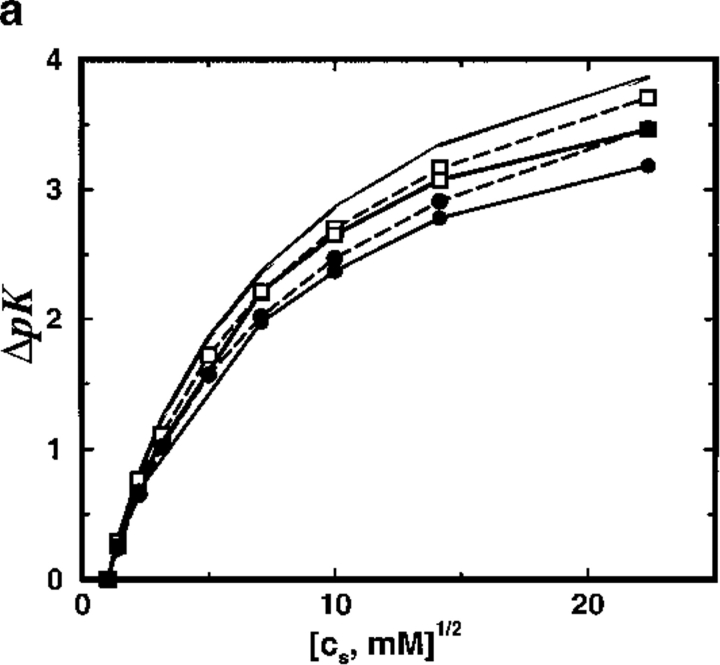
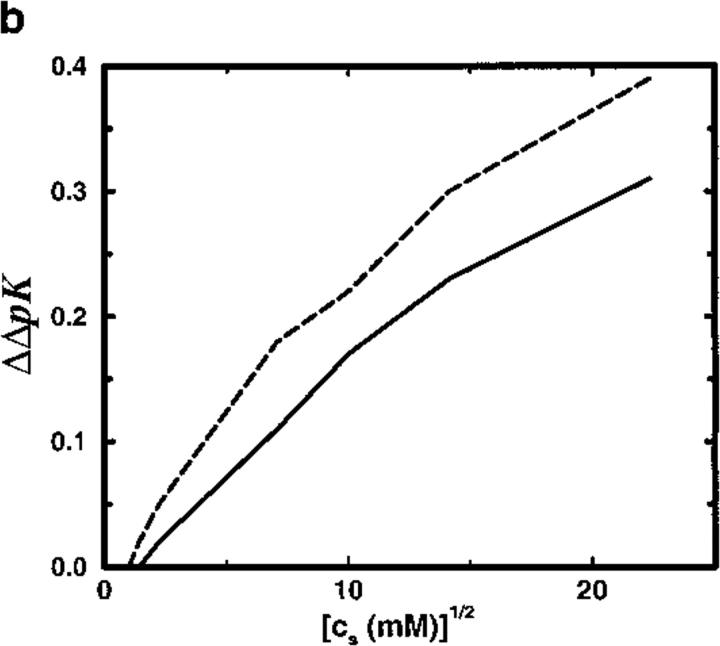
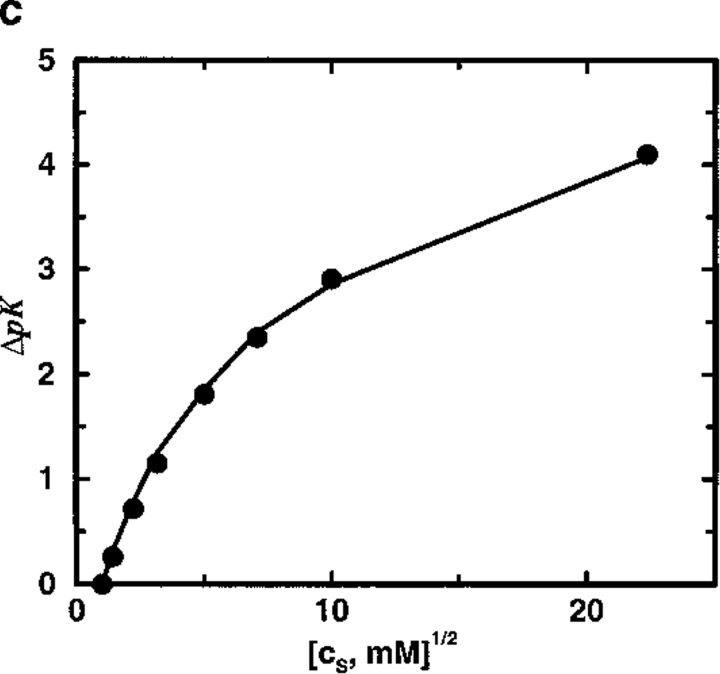
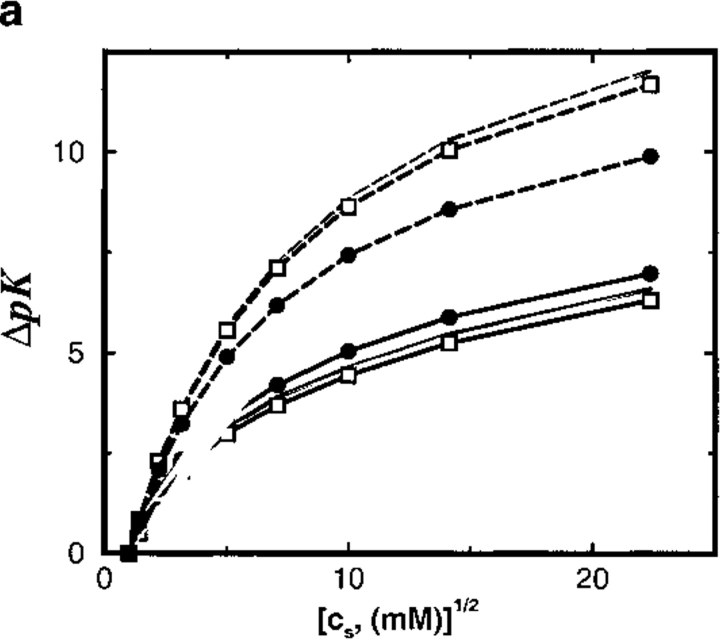
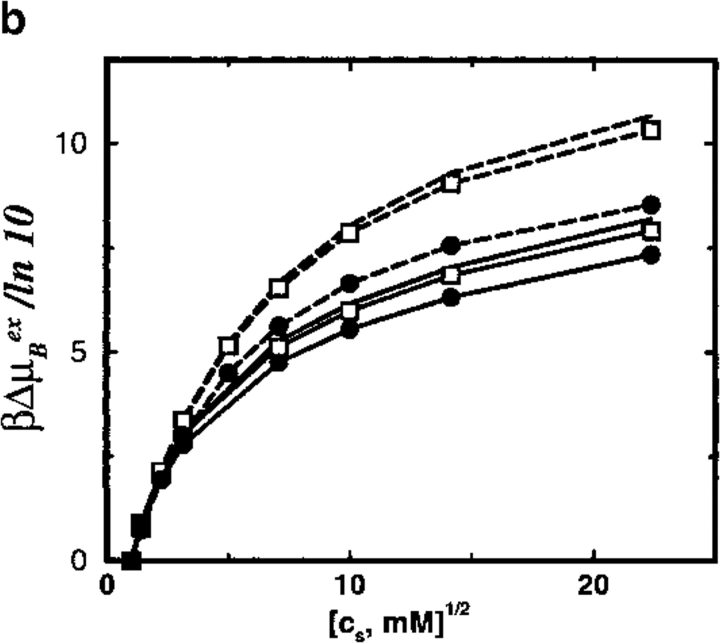
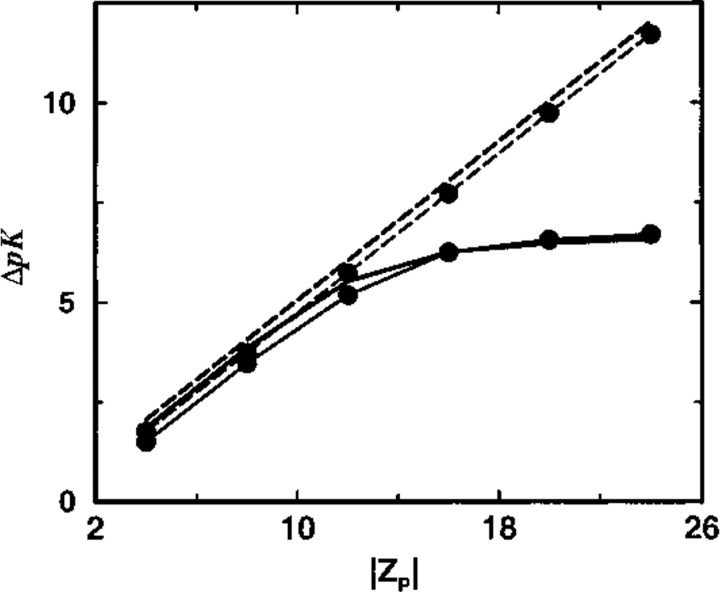
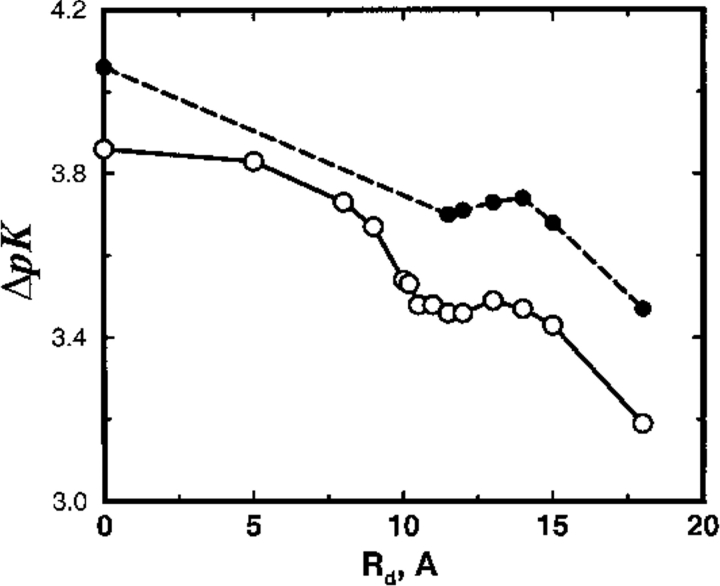
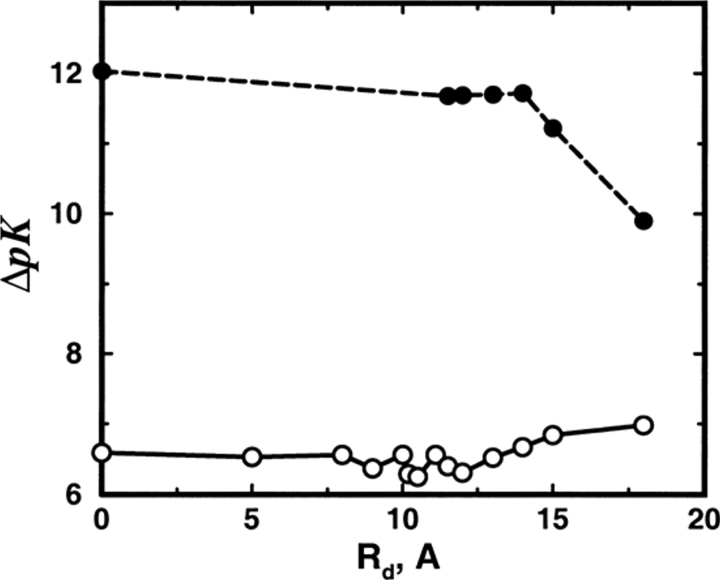
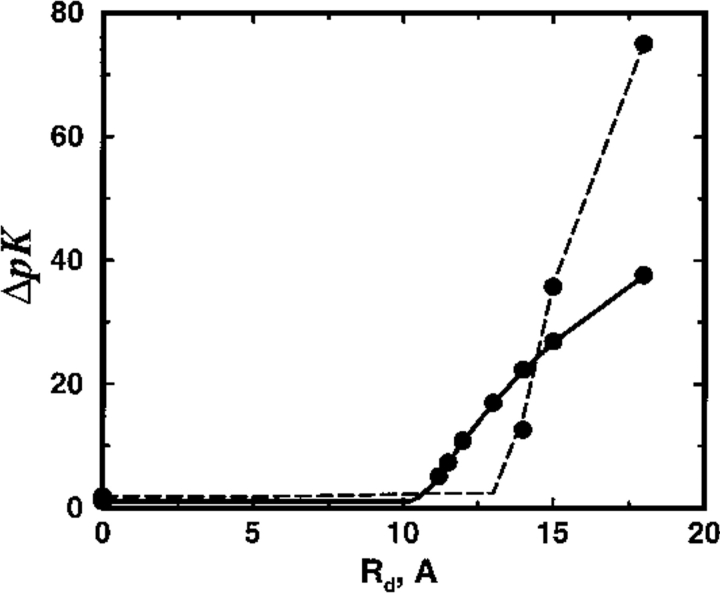
Monte Carlo simulations are used to assess the adequacy of the Tanford-Kirkwood prescription for electrostatic interactions in macromolecules. Within a continuum dielectric framework, the approach accurately describes salt screening of electrostatic interactions for moderately charged systems consistent with common proteins at physiological conditions. The limitations of the Debye-Hückel theory, which forms the statistical mechanical basis for the Tanford-Kirkwood result, become apparent for highly charged systems. It is shown, both by an analysis of the Debye-Hückel theory and by numerical simulations, that the difference in dielectric permittivity between macromolecule and surrounding solvent does not play a significant role for salt effects if the macromolecule is highly charged. By comparison to experimental data, the continuum dielectric model (combined with either an approximate effective Hamiltonian as in the Tanford-Kirkwood treatment or with exact Monte Carlo simulations) satisfactorily predicts the effects of charge mutation on metal ion binding constants, but only if the macromolecule and solvent are assigned the same or similar permittivities.
Figures
Similar articles
-
Monte Carlo and modified Tanford-Kirkwood results for macromolecular electrostatics calculations.J Phys Chem B. 2006 May 4;110(17):8832-9. doi: 10.1021/jp054891e. J Phys Chem B. 2006. PMID: 16640442
-
Tanford-Kirkwood electrostatics for protein modeling.Proc Natl Acad Sci U S A. 1999 Sep 28;96(20):11145-50. doi: 10.1073/pnas.96.20.11145. Proc Natl Acad Sci U S A. 1999. PMID: 10500144 Free PMC article.
-
Self-consistent field approach to protein structure and stability. I: pH dependence of electrostatic contribution.Proteins. 1997 Apr;27(4):576-96. doi: 10.1002/(sici)1097-0134(199704)27:4<576::aid-prot10>3.0.co;2-h. Proteins. 1997. PMID: 9141137
-
Protein electrostatics: a review of the equations and methods used to model electrostatic equations in biomolecules--applications in biotechnology.Biotechnol Annu Rev. 2003;9:315-95. doi: 10.1016/s1387-2656(03)09010-0. Biotechnol Annu Rev. 2003. PMID: 14650935 Review.
-
Development of constant-pH simulation methods in implicit solvent and applications in biomolecular systems.Biophys Rev. 2017 Oct;9(5):699-728. doi: 10.1007/s12551-017-0311-5. Epub 2017 Sep 18. Biophys Rev. 2017. PMID: 28921104 Free PMC article. Review.
Cited by
-
Coarse-grained dynamic RNA titration simulations.Interface Focus. 2019 Jun 6;9(3):20180066. doi: 10.1098/rsfs.2018.0066. Epub 2019 Apr 19. Interface Focus. 2019. PMID: 31065339 Free PMC article.
-
Calculating pH-dependent free energy of proteins by using Monte Carlo protonation probabilities of ionizable residues.Protein Cell. 2012 Mar;3(3):230-8. doi: 10.1007/s13238-012-2035-4. Epub 2012 Mar 31. Protein Cell. 2012. PMID: 22467263 Free PMC article.
-
pH Dependence of Charge Multipole Moments in Proteins.Biophys J. 2017 Oct 3;113(7):1454-1465. doi: 10.1016/j.bpj.2017.08.017. Biophys J. 2017. PMID: 28978439 Free PMC article.
-
Salt enhances calmodulin-target interaction.Biophys J. 2006 Apr 15;90(8):2903-10. doi: 10.1529/biophysj.105.068718. Epub 2006 Jan 20. Biophys J. 2006. PMID: 16428276 Free PMC article.
-
Sulfate anion stabilization of native ribonuclease A both by anion binding and by the Hofmeister effect.Protein Sci. 2002 Jul;11(7):1771-8. doi: 10.1110/ps.0205902. Protein Sci. 2002. PMID: 12070329 Free PMC article.
References
-
- Antonsiewicz, J., McCammon, J.A., and Gilson, M.K. 1994a. The determinants of pKas in proteins. Biochemistry 35 7819–7833. - PubMed
-
- Antonsiewicz, J., McCammon, J.A., and Gilson, M.K. 1994b. Prediction of pH-dependence properties of proteins. J. Mol. Biol. 238 415–436. - PubMed
-
- Bashford, D. and Gerwert, K. 1992. Electrostatic calculations of the pKa values of ionizable groups in bacteriorhodopsin. J. Mol. Biol. 224 473–486. - PubMed
-
- Bashford, D. and Karplus, M. 1990. pKa's of ionizable groups in proteins: Atomic detail from a continuum electrostatic model. Biochemistry 29 10219–10225. - PubMed
-
- Bashford, D., Karplus, M., and Canters, G.W. 1988. Electrostatic effects of charge pertubations introduced by metal oxidation in proteins—a theoretical analysis. J. Mol. Biol. 203 507–510. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
