

Review
doi: 10.1021/bi0155091.
Advances in determination of a high-resolution three-dimensional structure of rhodopsin, a model of G-protein-coupled receptors (GPCRs)
Affiliations
- PMID: 11425302
- PMCID: PMC1698954
- DOI: 10.1021/bi0155091
Item in Clipboard
Review
Advances in determination of a high-resolution three-dimensional structure of rhodopsin, a model of G-protein-coupled receptors (GPCRs)
Biochemistry.
.
No abstract available
Figures
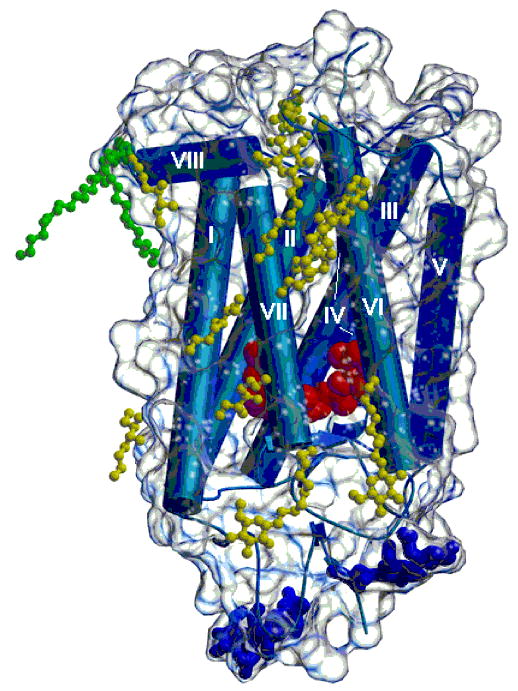
Three-dimensional crystal structure of rhodopsin with bound detergent and amphiphile molecules. Helical portions of the protein, including the seven transmembrane helices, are shown as blue rods, and β-strands are shown as blue arrows. The polypeptide connecting the helices appears as blue coils. A transparent envelope around the protein represents the molecular surface. The dark blue ball-and-stick groups at the bottom of the figure denote carbohydrate groups attached to the protein. Two palmitoyl groups covalently attached to the protein are shown in green. Nonylglucoside and heptanetriol molecules located near the hydrophobic surface of the protein are shown in yellow. The figure was drawn using Molscript (116) and Raster3d (117).
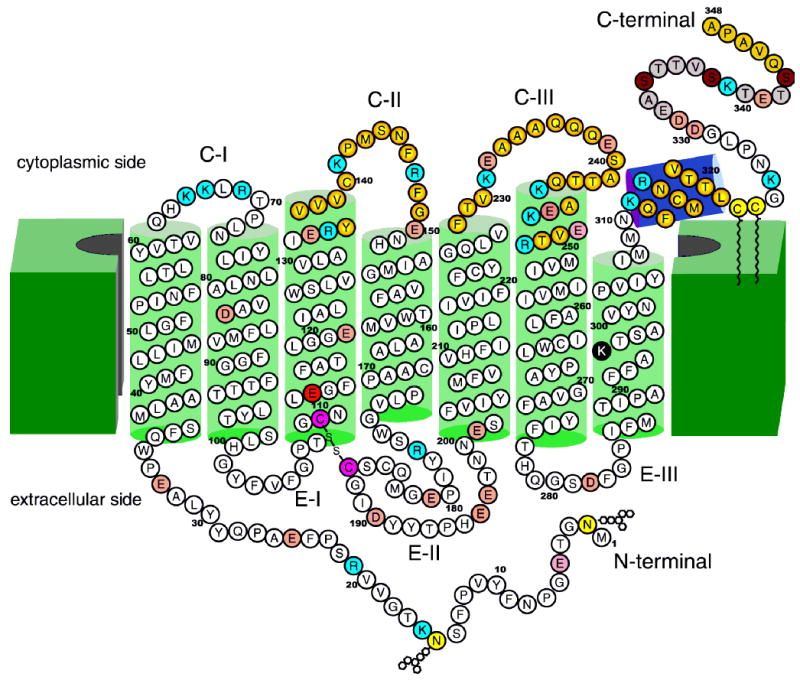
Two-dimensional model of bovine rhodopsin adopted after that of Hargrave (9). About ¼ of the polypeptide chain of opsins has cytoplasm exposure; ¼ is sequestered in the intradiscal space, and ½ forms a core of transmembrane helices. Green cylinders represent helices that in large part are imbedded in the disk membranes (19–34 residues each). The violet cylinder represents the amphiphilic helix (H-VIII) that runs parallel to the membranes. The color of rhodopsin results from the protonated Schiff base linkage of the 11-cis-retinal chromophore (100). Lys296 forms this protonated Schiff base with retinal and is shown as a black filled circle. Glu113 is a counterion of the Schiff base and is shown as a red filled circle. The disulfide bridge conserved among GPCRs is shown as purple filled circles (60). Two carbohydrate moieties at Asn2 and Asp15 and two palmitoyl groups at Cys322 and Cys323 (15, 64–71) are shown as light yellow filled circles (32, 72, 73). Light red filled circles represent acidic residues, and blue filled circles represent basic residues. Photoisomerization of the 11-cis-retinal chromophore of rhodopsin to all-trans-retinal leads to a conformational change in the protein, including the cytoplasmic surface (9–14, 32, 118, 119) leading to activation of Gt. Loops CII and CIII and helix VIII are involved in the recognition of Gt and are marked with filled yellow circles at the center, changing to red filled circles in the periphery. These residues are also important in binding of two regulatory proteins: rhodopsin kinase and arrestin. The functional phosphorylation occurs in the C-terminal region (represented by light brown filled circles) at phosphorylation sites denoted by dark brown filled circles (86–91, 96). The last five C-terminal residues were postulated to be involved in the vectorial transport within highly differentiated rod photoreceptors (92, 93, 120). The carbohydrate chains and the disulfide linkage (in pink filled circles) are oriented toward the lumen (extracellular) face of rhodopsin, and the C-terminal domain is cytoplasmic. To date, more than 100 mutations in the human rhodopsin gene have been associated with recessive and dominant retinitis pigmentosa (RP), as well as congenital stationary night blindness (CSNB).
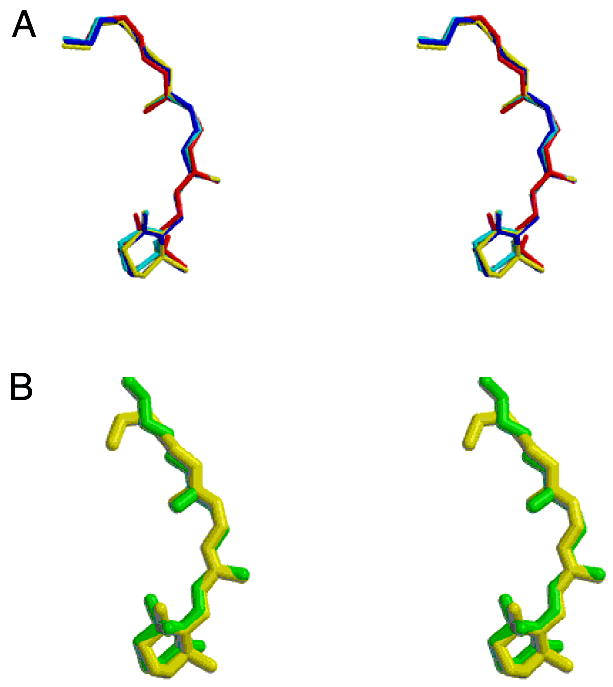
Conformation of the retinylidine chromophore. (A) Stereoview of the chromophores from the refined rhodopsin models showing the small variation in the conformation and orientation of the group in the four refined subunits. The rms deviation among the four molecules is 0.26 Å for all atoms. (B) Stereoview showing the chromophore from molecule A of the crystallographic model refined against twinned data (yellow) and the chromophore structure obtained by Watts and colleagues (108) (green).
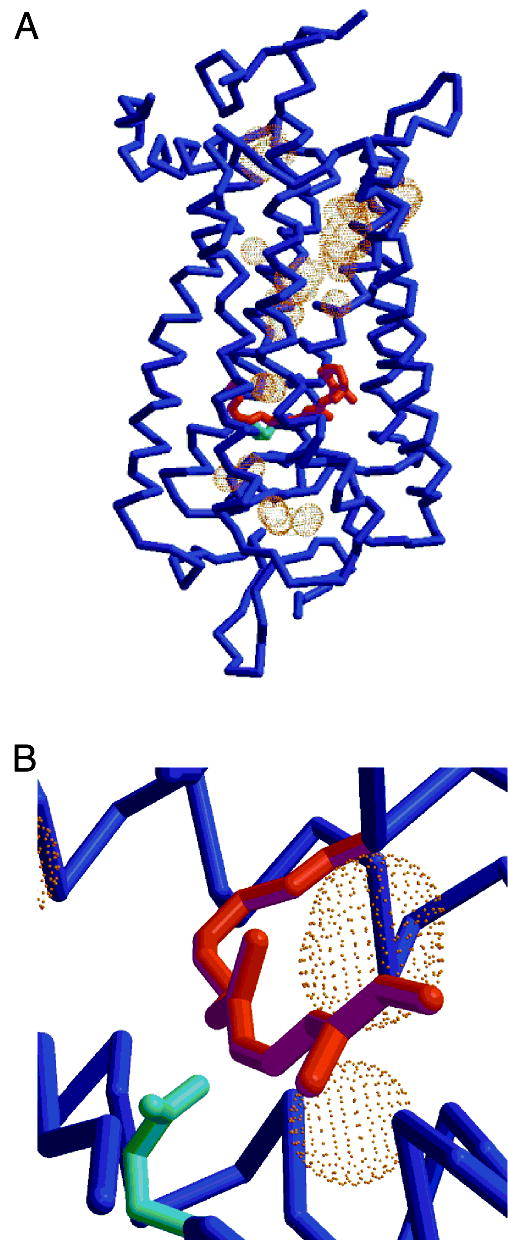
Cavities within the rhodopsin molecule. (A) Cα tracing (blue) with cavities shown by molecular surface dots around the test probes (1.4 Å radius). Glu113 is depicted in cyan and is about 3.6 Å from the Lys296 NZ. The cavities are of sufficient size to hold one or more water molecules. (B) Closeup of two cavities adjacent to the protonated Schiff base portion of the chromophore.
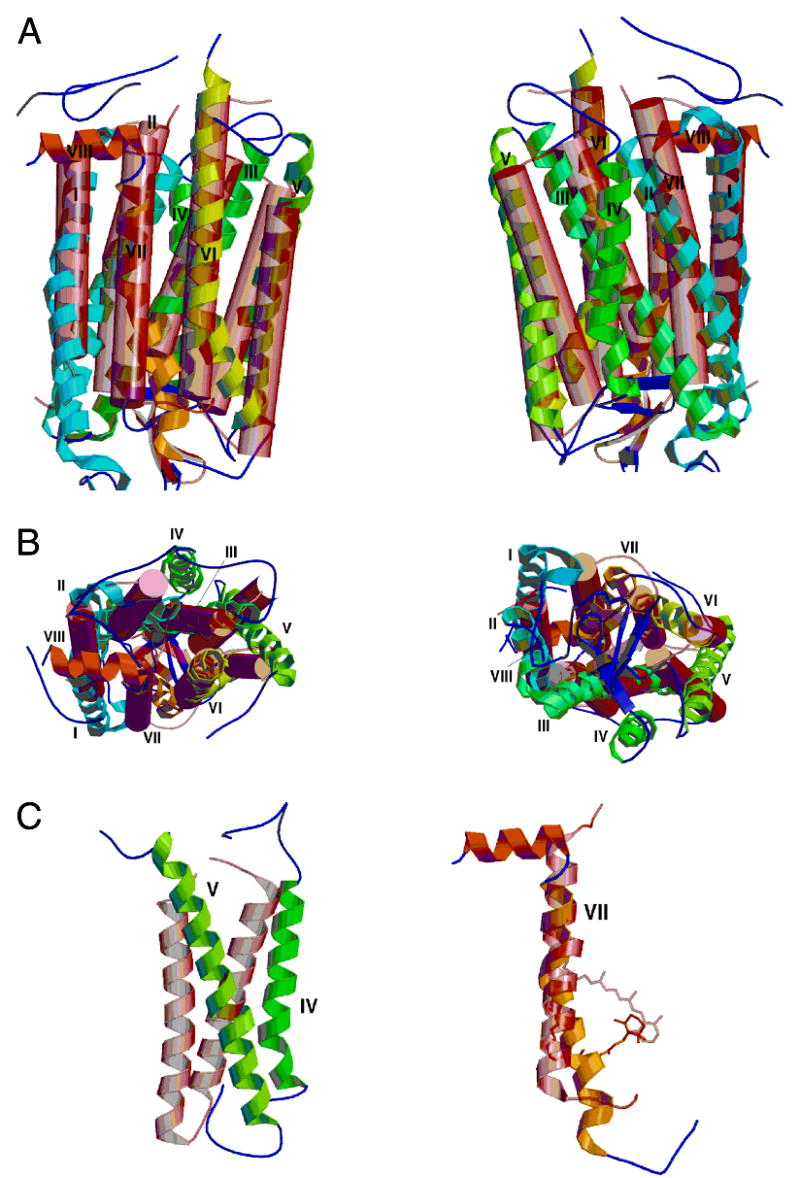
Superposition of bacteriorhodopsin (pink transparent cylinders and connecting coil) on molecule A of rhodopsin (colored helical ribbons and connecting coils). (A) At the left are molecules in the same orientation as in Figure 1. At the right is shown a similar view, with the molecules rotated 180° about the vertical axis. (B) At the left is a view of the top surface of the molecules. This is the cytoplasmic surface of rhodopsin. Note the substantial differences between helices IV and V in the two molecules. At the right is a bottom view of the molecules. (C) At the left, helices IV and V in the two molecules do not overlap significantly. At the right is helix VII. Note the irregular and kinked helix in rhodopsin as well as the differences in the location of the Schiff base attachment and orientation of the retinal chromophore.
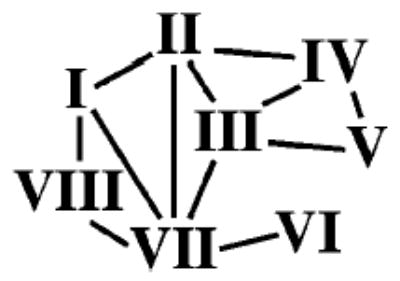
Connections between Helices via Hydrogen Bondinga a The helix–helix hydrogen bond interactions parallel the loss of accessible surface.
References
-
- Kuhlbrandt W, Gouaux E. Curr Opin Struct Biol. 1999;9:445–7. - PubMed
-
- Caffrey M. Curr Opin Struct Biol. 2000;10:486–97. - PubMed
-
- Adler E, Hoon MA, Mueller KL, Chandrashekar J, Ryba NJ, Zuker CS. Cell. 2000;100:693–702. - PubMed
-
- Mombaerts P. Science. 1999;286:707–11. - PubMed
-
- Milligan G, Rees S. Trends Pharmacol Sci. 1999;20:118–24. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases