

Neuronal cyclic AMP controls the developmental loss in ability of axons to regenerate
- PMID: 11425900
- PMCID: PMC6762375
- DOI: 10.1523/JNEUROSCI.21-13-04731.2001
Neuronal cyclic AMP controls the developmental loss in ability of axons to regenerate
Abstract
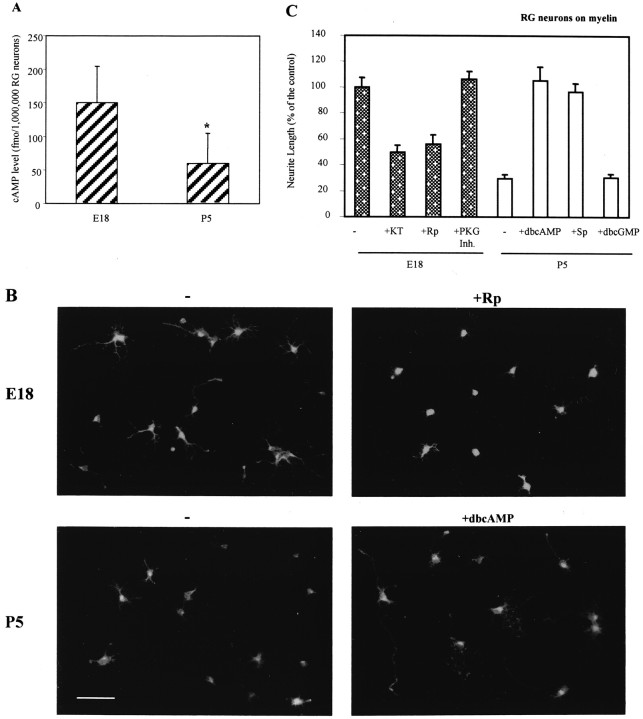
Unlike neonatal axons, mammalian adult axons do not regenerate after injury. Likewise, myelin, a major factor in preventing regeneration in the adult, inhibits regeneration from older but not younger neurons. Identification of the molecular events responsible for this developmental loss of regenerative capacity is believed key to devising strategies to encourage regeneration in adults after injury. Here, we report that the endogenous levels of the cyclic nucleotide, cAMP, are dramatically higher in young neurons in which axonal growth is promoted both by myelin in general and by a specific myelin component, myelin-associated glycoprotein (MAG), than in the same types of neurons that, when older, are inhibited by myelin-MAG. Inhibiting a downstream effector of cAMP [protein kinase A (PKA)] prevents myelin-MAG promotion from young neurons, and elevating cAMP blocks myelin-MAG inhibition of neurite outgrowth in older neurons. Importantly, developmental plasticity of spinal tract axons in neonatal rat pups in vivo is dramatically reduced by inhibition of PKA. Thus, the switch from promotion to inhibition by myelin-MAG, which marks the developmental loss of regenerative capacity, is mediated by a developmentally regulated decrease in endogenous neuronal cAMP levels.
Figures





Similar articles
-
Regeneration in the central nervous system.Exp Gerontol. 2003 Jan-Feb;38(1-2):79-86. doi: 10.1016/s0531-5565(02)00165-1. Exp Gerontol. 2003. PMID: 12543264
-
Prior exposure to neurotrophins blocks inhibition of axonal regeneration by MAG and myelin via a cAMP-dependent mechanism.Neuron. 1999 Jan;22(1):89-101. doi: 10.1016/s0896-6273(00)80681-9. Neuron. 1999. PMID: 10027292
-
Complement protein C1q modulates neurite outgrowth in vitro and spinal cord axon regeneration in vivo.J Neurosci. 2015 Mar 11;35(10):4332-49. doi: 10.1523/JNEUROSCI.4473-12.2015. J Neurosci. 2015. PMID: 25762679 Free PMC article.
-
Overcoming inhibitors in myelin to promote axonal regeneration.J Neurol Sci. 2005 Jun 15;233(1-2):43-7. doi: 10.1016/j.jns.2005.03.023. Epub 2005 Apr 20. J Neurol Sci. 2005. PMID: 15949495 Review.
-
Glial inhibition of nerve regeneration in the mature mammalian CNS.Glia. 2000 Jan 15;29(2):166-74. Glia. 2000. PMID: 10625335 Review.
Cited by
-
Ligand-Induced GPR110 Activation Facilitates Axon Growth after Injury.Int J Mol Sci. 2021 Mar 25;22(7):3386. doi: 10.3390/ijms22073386. Int J Mol Sci. 2021. PMID: 33806166 Free PMC article.
-
Association of VSNL1 with schizophrenia, frontal cortical function, and biological significance for its gene product as a modulator of cAMP levels and neuronal morphology.Transl Psychiatry. 2011 Jul 19;1(7):e22. doi: 10.1038/tp.2011.20. Transl Psychiatry. 2011. PMID: 22832524 Free PMC article.
-
Role of NG2 in development and regeneration.J Neurosci. 2006 Jul 5;26(27):7127-8. doi: 10.1523/jneurosci.2104-06.2006. J Neurosci. 2006. PMID: 16826634 Free PMC article. Review. No abstract available.
-
EphB3: an endogenous mediator of adult axonal plasticity and regrowth after CNS injury.J Neurosci. 2006 Mar 22;26(12):3087-101. doi: 10.1523/JNEUROSCI.4797-05.2006. J Neurosci. 2006. PMID: 16554460 Free PMC article.
-
Protein kinase A activation promotes plasma membrane insertion of DCC from an intracellular pool: A novel mechanism regulating commissural axon extension.J Neurosci. 2004 Mar 24;24(12):3040-50. doi: 10.1523/JNEUROSCI.4934-03.2004. J Neurosci. 2004. PMID: 15044543 Free PMC article.
References
-
- Bashaw GJ, Goodman CS. Chimeric axon guidance receptors: the cytoplasmic domains of slit and netrin receptors specify attraction versus repulsion. Cell. 1999;97:917–926. - PubMed
-
- Bates CA, Stelzner DJ. Extension and regeneration of corticospinal axons after early spinal injury and the maintenance of corticospinal topography. Exp Neurol. 1993;123:106–117. - PubMed
-
- Bernstein-Goral H, Bregman BS. Spinal cord transplants support the regeneration of axotomized neurons after spinal cord lesions at birth: a quantitative double-labeling study. Exp Neurol. 1993;123:118–132. - PubMed
-
- Bregman BS. Spinal cord transplants permit the growth of serotonergic axons across the site of neonatal spinal cord transection. Brain Res. 1987a;431:265–279. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous