

Induction of primordial germ cells from murine epiblasts by synergistic action of BMP4 and BMP8B signaling pathways
- PMID: 11427739
- PMCID: PMC35432
- DOI: 10.1073/pnas.151242798
Induction of primordial germ cells from murine epiblasts by synergistic action of BMP4 and BMP8B signaling pathways
Abstract
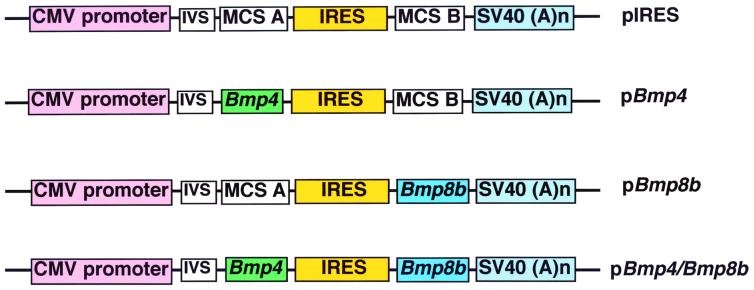
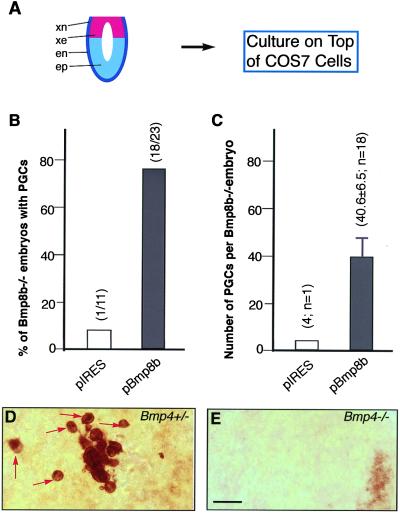
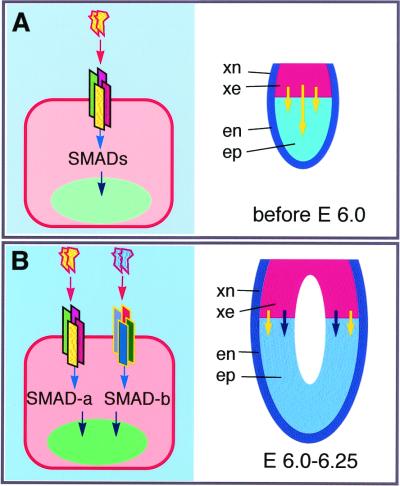
Extraembryonic ectoderm-derived factors instruct the pluripotent epiblast cells to develop toward a restricted primordial germ cell (PGC) fate during murine gastrulation. Genes encoding Bmp4 of the Dpp class and Bmp8b of the 60A class are expressed in the extraembryonic ectoderm and targeted mutation of either results in severe defects in PGC formation. It has been shown that heterodimers of DPP and 60A classes of bone morphogenetic proteins (BMPs) are more potent than each homodimers in bone and mesoderm induction in vitro, suggesting that BMP4 and BMP8B may form heterodimers to induce PGCs. To investigate how BMP4 and BMP8B interact and signal for PGC induction, we cocultured epiblasts of embryonic day 6.0--6.25 embryos with BMP4 and BMP8B proteins produced by COS cells. Our data show that BMP4 or BMP8B homodimers alone cannot induce PGCs whereas they can in combination, providing evidence that two BMP pathways are simultaneously required for the generation of a given cell type in mammals and also providing a prototype method for PGC induction in vitro. Furthermore, the PGC defects of Bmp8b mutants can be rescued by BMP8B homodimers whereas BMP4 homodimers cannot mitigate the PGC defects of Bmp4 null mutants, suggesting that BMP4 proteins are also required for epiblast cells to gain germ-line competency before the synergistic action of BMP4 and BMP8B.
Figures


References
-
- Eddy E M. Int Rev Cytol. 1975;43:229–280. - PubMed
-
- Wylie C. Cell. 1999;96:165–174. - PubMed
-
- McLaren A. Genes Dev. 1999;13:373–376. - PubMed
-
- Weidinger G, Wolke U, Koprunner M, Klinger M, Raz E. Development (Cambridge, UK) 1999;126:5295–5307. - PubMed
-
- Kloc M, Bilinski S, Chan A P, Allen L H, Zearfoss N R, Etkin L D. Int Rev Cytol. 2001;203:63–91. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
