

PTP-PEST, a scaffold protein tyrosine phosphatase, negatively regulates lymphocyte activation by targeting a unique set of substrates
- PMID: 11432829
- PMCID: PMC125513
- DOI: 10.1093/emboj/20.13.3414
PTP-PEST, a scaffold protein tyrosine phosphatase, negatively regulates lymphocyte activation by targeting a unique set of substrates
Abstract
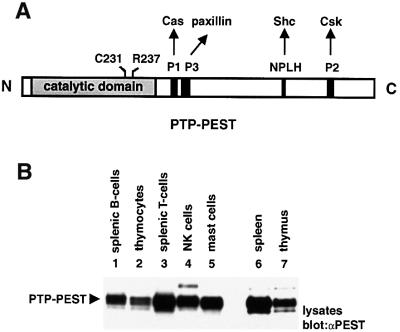
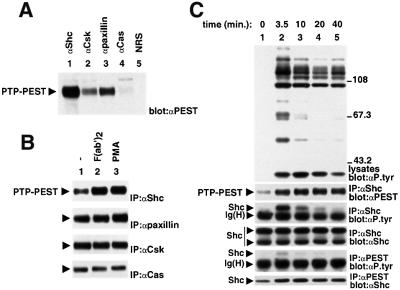
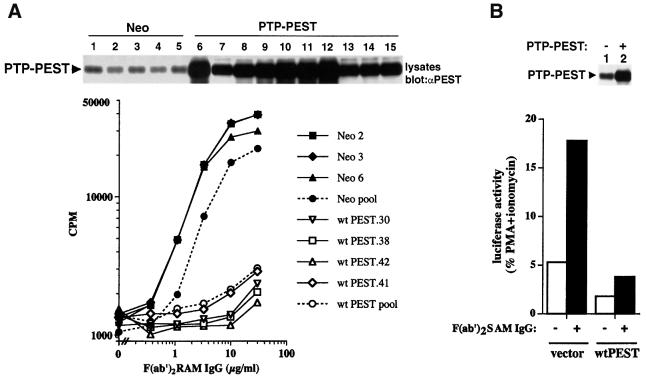
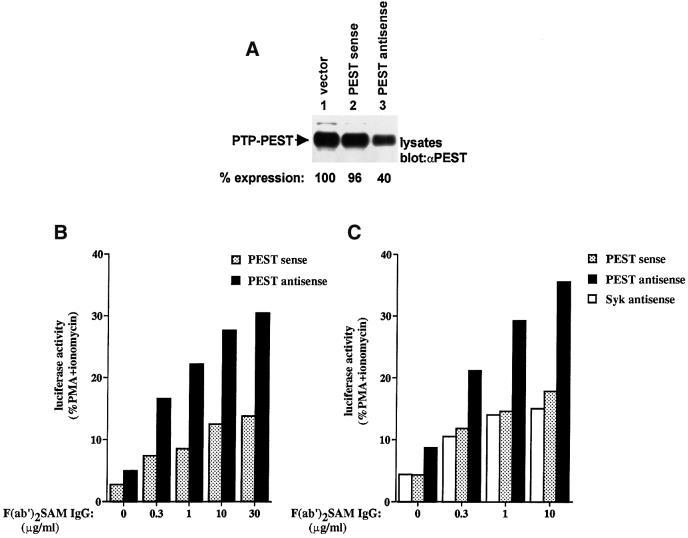
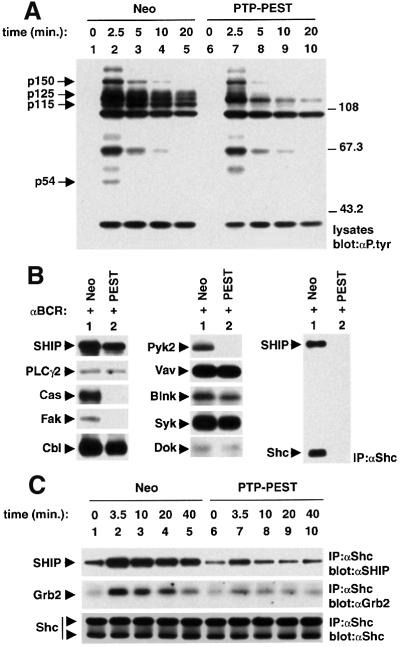
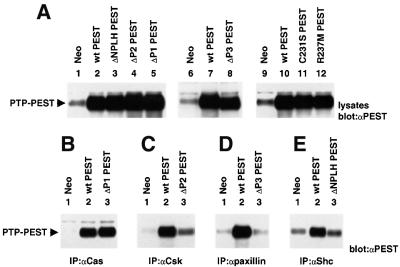
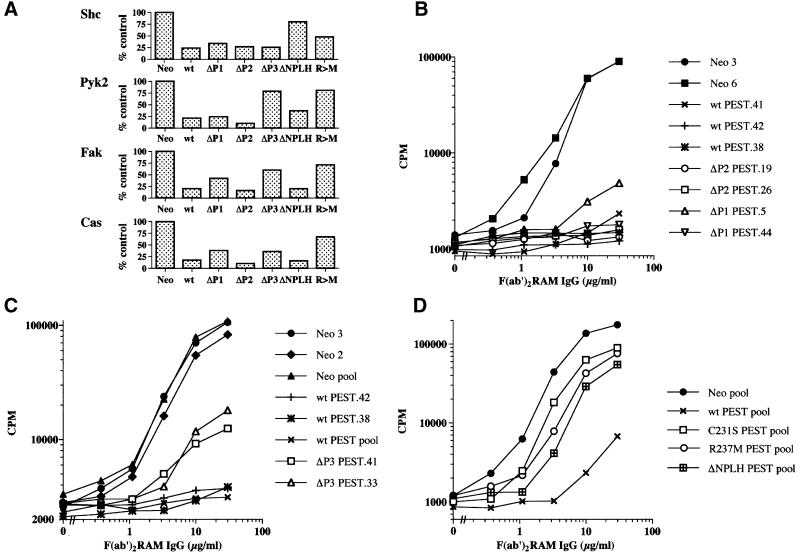
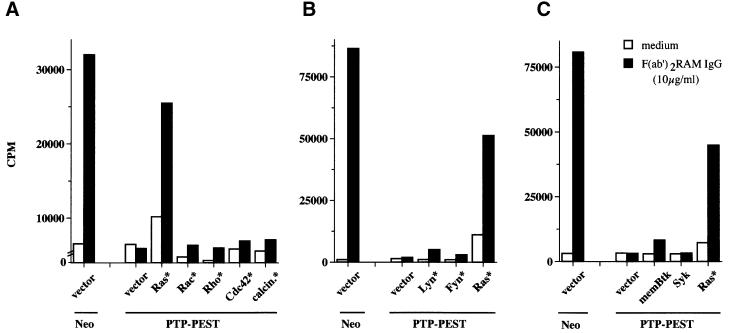
There is increasing interest in elucidating the mechanisms involved in the negative regulation of lymphocyte activation. Herein, we show that the cytosolic protein tyrosine phosphatase PTP-PEST is expressed abundantly in a wide variety of haemopoietic cell types, including B cells and T cells. In a model B-cell line, PTP-PEST was found to be constitutively associated with several signalling molecules, including Shc, paxillin, Csk and Cas. The interaction between Shc and PTP-PEST was augmented further by antigen receptor stimulation. Overexpression studies, antisense experiments and structure-function analyses provided evidence that PTP-PEST is an efficient negative regulator of lymphocyte activation. This function correlated with the ability of PTP-PEST to induce dephosphorylation of Shc, Pyk2, Fak and Cas, and inactivate the Ras pathway. Taken together, these data suggest that PTP-PEST is a novel and unique component of the inhibitory signalling machinery in lymphocytes.
Figures
Similar articles
-
Intact LIM 3 and LIM 4 domains of paxillin are required for the association to a novel polyproline region (Pro 2) of protein-tyrosine phosphatase-PEST.J Biol Chem. 1999 Jul 16;274(29):20550-60. doi: 10.1074/jbc.274.29.20550. J Biol Chem. 1999. PMID: 10400685
-
Combination of gene targeting and substrate trapping to identify substrates of protein tyrosine phosphatases using PTP-PEST as a model.Biochemistry. 1998 Sep 22;37(38):13128-37. doi: 10.1021/bi981259l. Biochemistry. 1998. PMID: 9748319
-
Negative signaling in B cells causes reduced Ras activity by reducing Shc-Grb2 interactions.J Immunol. 1997 Feb 1;158(3):1125-32. J Immunol. 1997. PMID: 9013951
-
Adapter molecules in T cell receptor signaling.Inflamm Bowel Dis. 1999 May;5(2):107-18. doi: 10.1097/00054725-199905000-00007. Inflamm Bowel Dis. 1999. PMID: 10338380 Review. No abstract available.
-
Regulation of tumor cell migration by protein tyrosine phosphatase (PTP)-proline-, glutamate-, serine-,and threonine-rich sequence (PEST).Chin J Cancer. 2013 Feb;32(2):75-83. doi: 10.5732/cjc.012.10084. Epub 2012 Dec 7. Chin J Cancer. 2013. PMID: 23237212 Free PMC article. Review.
Cited by
-
Focal adhesion kinases in adhesion structures and disease.J Signal Transduct. 2012;2012:296450. doi: 10.1155/2012/296450. Epub 2012 Jul 19. J Signal Transduct. 2012. PMID: 22888421 Free PMC article.
-
High-level expression of protein tyrosine phosphatase non-receptor 12 is a strong and independent predictor of poor prognosis in prostate cancer.BMC Cancer. 2019 Oct 12;19(1):944. doi: 10.1186/s12885-019-6182-3. BMC Cancer. 2019. PMID: 31606028 Free PMC article.
-
Analysis of the rs2476601 polymorphism of PTPN22 in Mexican mestizo patients with leprosy.Biomed Rep. 2019 Feb;10(2):127-132. doi: 10.3892/br.2019.1184. Epub 2019 Jan 7. Biomed Rep. 2019. PMID: 30675352 Free PMC article.
-
Comparison of T cell receptor-induced proximal signaling and downstream functions in immortalized and primary T cells.PLoS One. 2009;4(5):e5430. doi: 10.1371/journal.pone.0005430. Epub 2009 May 4. PLoS One. 2009. PMID: 19412549 Free PMC article.
-
Leupaxin Expression Is Dispensable for B Cell Immune Responses.Front Immunol. 2020 Mar 25;11:466. doi: 10.3389/fimmu.2020.00466. eCollection 2020. Front Immunol. 2020. PMID: 32269569 Free PMC article.
References
-
- Benschop R.J. and Cambier,J.C. (1999) B cell development: signal transduction by antigen receptors and their surrogates. Curr. Opin. Immunol., 11, 143–151. - PubMed
-
- Bolland S., Pearse,R.N., Kurosaki,T. and Ravetch,J.V. (1998) SHIP modulates immune receptor responses by regulating membrane association of Btk. Immunity, 8, 509–516. - PubMed
-
- Campbell K.S. (1999) Signal transduction from the B cell antigen-receptor. Curr. Opin. Immunol., 11, 256–264. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
