

doi: 10.1093/emboj/20.13.3518.
An upstream repressor element plays a role in Igf2 imprinting
Affiliations
- PMID: 11432838
- PMCID: PMC125515
- DOI: 10.1093/emboj/20.13.3518
Item in Clipboard
An upstream repressor element plays a role in Igf2 imprinting
EMBO J.
.
Abstract
The imprinted Igf2 gene is associated with a small upstream region that is differentially methylated on the active paternal allele. We have identified a repressor element within this sequence and shown that repression is probably mediated through a trans- acting factor, GCF2. DNA methylation of this site abrogates both protein binding and repressor activity. Targeting experiments demonstrate that this element plays a role in the repression of the maternal Igf2 gene in vivo.
Figures
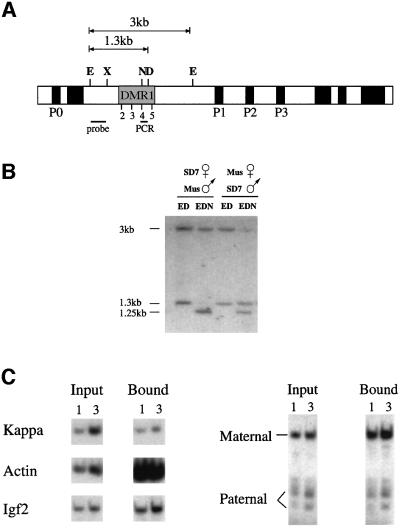
Fig. 1. Igf2 allelic methylation and histone acetylation patterns. (A) Map of the mouse Igf2 gene indicating the promoters (P0–P3), the differentially methylated (DMR1) region including HpaII sites 2–5, restriction sites for E (EcoRI), X (XbaI), N (NgoMI; NgoMIV), D (DraI; only present on the M.spretus allele), the EcoRI–XbaI probe and the position of the PCR fragment used in the immunoprecipitation experiments. (B) Igf2 differential methylation at site 4 was measured by Southern blot analysis of kidney genomic DNA from SD7 hetero zygous mice using the restriction enzymes shown in each lane. After digestion with NgoMI, both the domesticus allele (3 kb) and the spretus allele (1.3 kb) yield a 1.25 kb band. Similar results were obtained using DNA from liver but, in this case, the paternal allele was only 70% methylated. (C) The acetylation state of nucleosomes in DMR1 was determined by immunoprecipitation of mononucleosomes from SD7 heterozygous mice using anti-acetylated H4 histone. DNA from input and bound fractions was extracted and 1 or 3 µl subjected to quantitative radiolabeled PCR with primers specific for sequences from the κ-light chain, β-actin and Igf2 upstream regions (left panel). The Igf2 PCR product was digested with HpaII to identify a polymorphic site on this fragment, present only on the spretus (paternal) allele, analyzed by gel electrophoresis (right panel) and the results quantitated using a phosphoimager. The ratio between the maternal and paternal alleles in the bound fraction was found to be 5.5-fold more than that measured for the input DNA. In a separate experiment, we also determined that the PCR product obtained from total DNA is produced equally from the two alleles (data not shown).
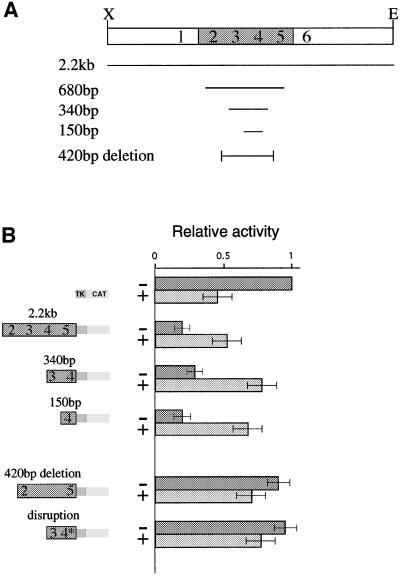
Fig. 2. DMR1 contains a methylation-sensitive repressor element. Transient transfection into 293 human kidney cells was used to measure repressor activity on a pTK-CAT reporter construct. (A) Map showing the different constructs used in the transient transfection experiments. (B) CAT assays of reporter constructs. The results are shown in graphic form relative to the basic TK-CAT vector. These values were determined by scanning autoradiograms. For each lane, the particular construct used is indicated by a modular diagram made up of fragments from DMR1, the TK promoter and the CAT reporter gene. The last construct (disruption) is the same as the 340 bp fragment but contains a loxP insertion (see text). Constructs were either unmethylated (–) or in vitro methylated by MHpaII (+). The HpaII sites (2–5) located within each fragment are indicated in the diagram. Most of these experiments were repeated 4–5 times and the results shown represent average values (± SD).
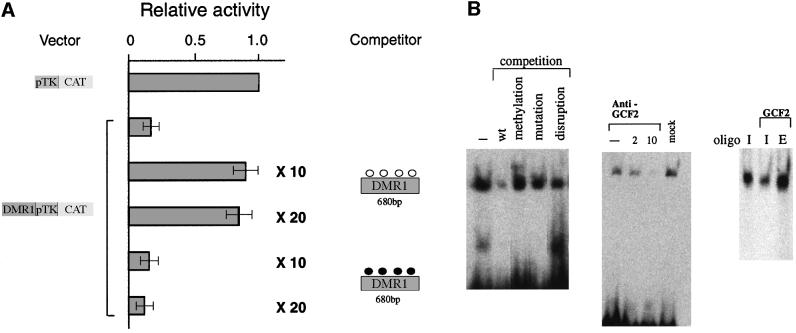
Fig. 3. Repression is mediated by protein interactions with DMR1. (A) Co-transfection competition experiments were carried out using a reporter construct containing a 680 bp fragment from DMR1 (see Figure 2A) and competitor plasmid DNA containing only the 680 bp fragment (10- or 20-fold excess in methylated or unmethylated form), and the results (average ± SD) are shown as in Figure 2. The pTk-CAT vector was unaffected by the addition of either unmethylated or methylated competitor DNA (data not shown). (B) Gel shift analysis of a 32P-end-labeled Igf2 upstream oligonucleotide using 293 cell nuclear extract. Competition (left) was carried out using a 45-fold excess of non-radioactive wild-type (wt), methylated or disrupted oligonucleotides (see Materials and methods). Gel shift analysis was also carried out (middle) with the same extract treated with two different concentrations of GCF2 antibody to bring about immunodepletion. In a separate experiment, we removed the antibody complex from the Sepharose beads and confirmed by western analysis that it contained GCF2. A mock experiment in which the extract was passed over protein A–Sepharose beads without antibody is also shown. Extract or pure recombinant GCF2 protein was reacted (right) with either the labeled Igf2 upstream oligonucleotide (I) or an oligonucleotide containing the EGFR recognition sequence (E). A small amount of immunodepleted extract was added to the pure protein in order to stabilize it and enhance binding.
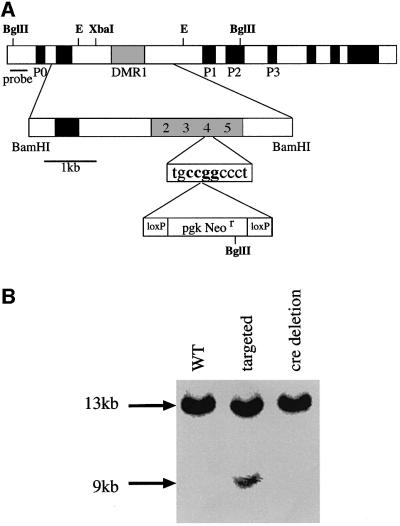
Fig. 4. Genomic targeting of a loxP element into DMR1 HpaII site 4. (A) The targeting construct. A 5 kb BamHI fragment containing DMR1 was subcloned from a genomic library and a loxP-Neor-loxP cassette (bottom line) was then inserted into HpaII site 4, which resides within a GCF2 consensus sequence (see Kageyama and Pastan, 1989; Reed et al., 1998). (B) Southern blot analysis of genomic DNA from wild-type (WT), targeted and cre-deleted ES cells restricted with BglII and probed with the fragments indicated in the map. Only data from the left hand probe are shown in the blot. The targeted allele appears under-represented in this figure because of the presence of DNA from accompanying feeder cells. The nature of the disruption was verified by amplifying and sequencing this region from the cre-deleted mice.
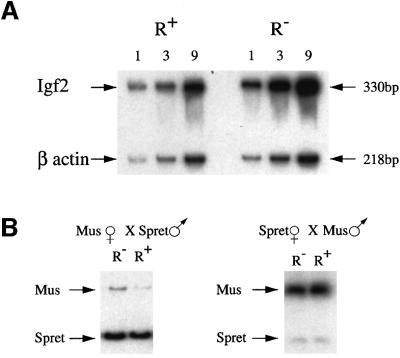
Fig. 5. Analysis of Igf2 maternal allele expression in targeted mice. (A) RT–PCR on total RNA extracted from 8–10 whole 12.5 d.p.c. embryos, which were first genotyped by PCR in order to identify and group the R+/Igf2– and R–/Igf2– individuals. PCR was carried out on 1, 3 or 9 µl of cDNA in the presence of [α-32P]dCTP, and the products were run on a 5% acrylamide gel. PCR for β-actin cDNA was carried out in the same tube and served as a control. Igf2 transcription in R– embryos was 2.9-fold higher than in R+ embryos as determined by phosphoimager analysis, after normalization for β-actin. Similar results were also obtained using pooled embryos from an additional inde pendent targeted mouse line. We also carried out expression analysis on individual embryos. In this case, the level of Igf2 mRNA was normalized against β-actin, and the average relative amount of Igf2 in the R+ embryos set at 1 (R+, 1.0 ± 0.2, n = 5; R–, 2.8 ± 0.6, n = 6, P <0.001). (B) RT–PCR was carried out on total RNA from 8–10 whole 12.5 d.p.c. embryos obtained from crosses between M.musculus mice carrying one targeted Igf2 allele (R–) and homozygous SD7 mice (R+). The PCR product was then cut with BsaAI, which recognizes a polymorphic site present only in the M.spretus allele, and was run on a 7% acrylamide gel. In the left panel, the R– allele is derived from the mother while, in the right panel, this allele is paternally derived. The R– maternal allele is 2.8-fold more active than the R+ allele when compared with the spretus paternal allele, as determined by phosphoimager analysis.
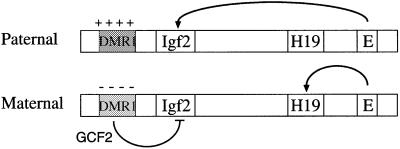
Fig. 6. Model of the Igf2 imprinting mechanism. The diagram shows the maternal and paternal Igf2/H19 alleles with the DMR1 methylated and packaged into a closed chromatin structure containing deacetylated histones on the paternal copy, while the DMR1 on the maternal copy is unmethylated and packaged into a more open structure containing acetylated histones. Enhancer sequences (E) are located 3′ to the H19 gene. On the maternal allele, these sequences activate H19, and Igf2 activity is repressed by elements in DMR1 that bind the GCF2 protein. On the paternal allele, DMR1-mediated repression is abrogated either because enhancer sequences overcome this inhibitory element or because of a closed methylated chromatin structure.
Similar articles
-
Epigenetic regulation of Igf2/H19 imprinting at CTCF insulator binding sites.J Cell Biochem. 2003 Dec 1;90(5):1038-55. doi: 10.1002/jcb.10684. J Cell Biochem. 2003. PMID: 14624463
-
Methylation of a CTCF-dependent boundary controls imprinted expression of the Igf2 gene.Nature. 2000 May 25;405(6785):482-5. doi: 10.1038/35013100. Nature. 2000. PMID: 10839546
-
CTCF binding sites promote transcription initiation and prevent DNA methylation on the maternal allele at the imprinted H19/Igf2 locus.Hum Mol Genet. 2006 Oct 1;15(19):2945-54. doi: 10.1093/hmg/ddl237. Epub 2006 Aug 23. Hum Mol Genet. 2006. PMID: 16928784
-
IVF results in de novo DNA methylation and histone methylation at an Igf2-H19 imprinting epigenetic switch.Mol Hum Reprod. 2005 Sep;11(9):631-40. doi: 10.1093/molehr/gah230. Epub 2005 Oct 11. Mol Hum Reprod. 2005. PMID: 16219628
-
Chromatin structure and imprinting: developmental control of DNase-I sensitivity in the mouse insulin-like growth factor 2 gene.Dev Genet. 1995;17(3):240-52. doi: 10.1002/dvg.1020170309. Dev Genet. 1995. PMID: 8565330
Cited by
-
DNA methylation and its role in the pathogenesis of diabetes.Pediatr Diabetes. 2017 May;18(3):167-177. doi: 10.1111/pedi.12521. Pediatr Diabetes. 2017. PMID: 28401680 Free PMC article. Review.
-
Callipyge mutation affects gene expression in cis: a potential role for chromatin structure.Genome Res. 2006 Mar;16(3):340-6. doi: 10.1101/gr.4389306. Epub 2006 Jan 13. Genome Res. 2006. PMID: 16415109 Free PMC article.
-
DNA methylation may restrict but does not determine differential gene expression at the Sgy/Tead2 locus during mouse development.Mol Cell Biol. 2004 Mar;24(5):1968-82. doi: 10.1128/MCB.24.5.1968-1982.2004. Mol Cell Biol. 2004. PMID: 14966277 Free PMC article.
-
Structural and functional analysis of a 0.5-Mb chicken region orthologous to the imprinted mammalian Ascl2/Mash2-Igf2-H19 region.Genome Res. 2005 Jan;15(1):154-65. doi: 10.1101/gr.2609605. Epub 2004 Dec 8. Genome Res. 2005. PMID: 15590938 Free PMC article.
-
Paradoxical role of DNA methylation in activation of FoxA2 gene expression during endoderm development.J Biol Chem. 2014 Aug 22;289(34):23882-92. doi: 10.1074/jbc.M114.573469. Epub 2014 Jul 11. J Biol Chem. 2014. PMID: 25016019 Free PMC article.
References
-
- Bartolomei M.S., Webber,A.L., Brunkow,M.E. and Tilghman,S.M. (1993) Epigenetic mechanisms underlying the imprinting of the mouse H19 gene. Genes Dev., 7, 1663–1673. - PubMed
-
- Borras T., Peterson,C.A. and Piatigorsky,J. (1988) Evidence for positive and negative regulation in the promoter of the chicken δ1-crystallin gene. Dev. Biol., 127, 209–219. - PubMed
-
- Constancia M., Pickard,B., Kelsey,G. and Reik,W. (1998) Imprinting mechanisms. Genome Res., 8, 881–900. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
