

Loss of poly(ADP-ribose) polymerase-1 causes increased tumour latency in p53-deficient mice
- PMID: 11432840
- PMCID: PMC125506
- DOI: 10.1093/emboj/20.13.3535
Loss of poly(ADP-ribose) polymerase-1 causes increased tumour latency in p53-deficient mice
Abstract
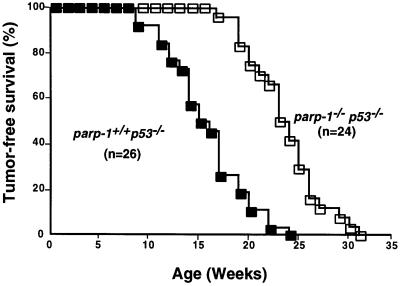
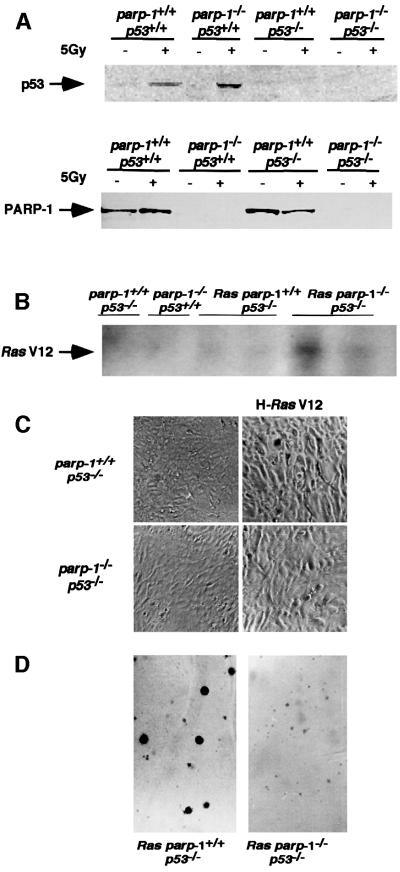
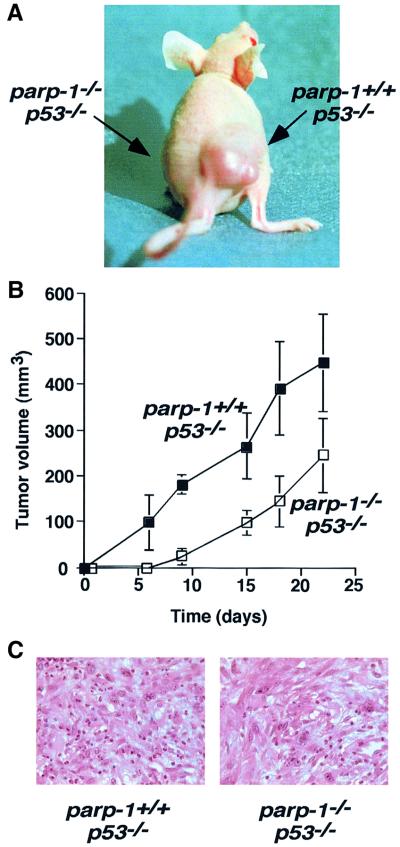
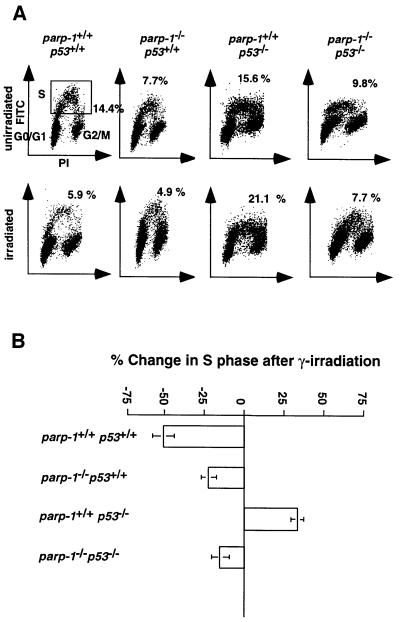
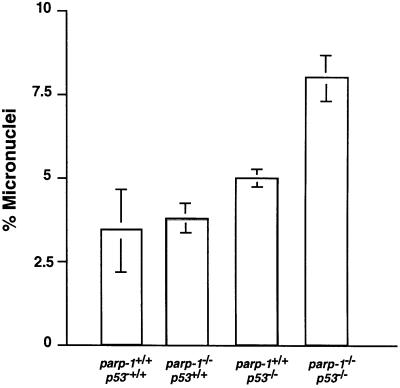
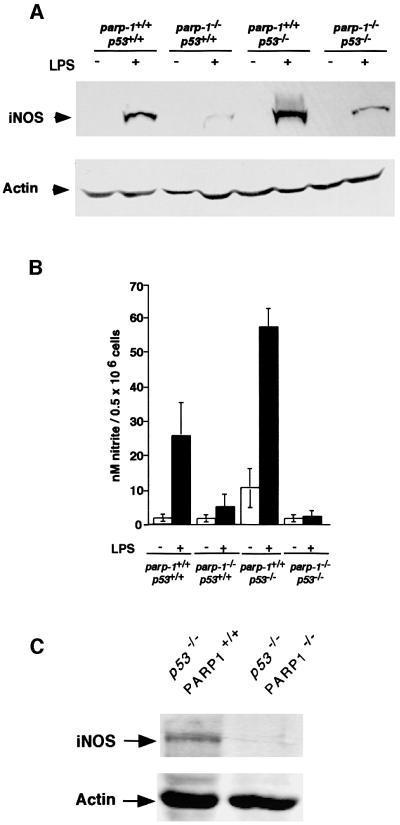
PARP-1-deficient mice display a severe defect in the base excision repair pathway leading to radiosensitivity and genomic instability. They are protected against necrosis induced by massive oxidative stress in various inflammatory processes. Mice lacking p53 are highly predisposed to malignancy resulting from defective cell cycle checkpoints, resistance to DNA damage-induced apoptosis as well as from upregulation of the iNOS gene resulting in chronic oxidative stress. Here, we report the generation of doubly null mutant mice. We found that tumour-free survival of parp-1(-/-)p53(-/-) mice increased by 50% compared with that of parp- 1(+/+)p53(-/-) mice. Tumour formation in nude mice injected with oncogenic parp-1(-/-)p53(-/-) fibroblasts was significantly delayed compared with parp-1(+/+)p53(-/-) cells. Upon gamma-irradiation, a partial restoration of S-phase radiosensitivity was found in parp-1(-/-)p53(-/-) primary fibroblasts compared with parp-1(+/+)p53(-/-) cells. In addition, iNOS expression and nitrite release were dramatically reduced in the parp-1(-/-)p53(-/-) mice compared with parp-1(+/+)p53(-/-) mice. The abrogation of the oxydated status of p53(-/-) cells, due to the absence of parp-1, may be the cause of the delay in the onset of tumorigenesis in parp-1(-/-)p53(-/-) mice.
Figures
Similar articles
-
DNA strand break-sensing molecule poly(ADP-Ribose) polymerase cooperates with p53 in telomere function, chromosome stability, and tumor suppression.Mol Cell Biol. 2001 Jun;21(12):4046-54. doi: 10.1128/MCB.21.12.4046-4054.2001. Mol Cell Biol. 2001. PMID: 11359911 Free PMC article.
-
Poly(ADP-ribose) polymerase inhibitors activate the p53 signaling pathway in neural stem/progenitor cells.BMC Neurosci. 2017 Jan 17;18(1):14. doi: 10.1186/s12868-016-0333-0. BMC Neurosci. 2017. PMID: 28095779 Free PMC article.
-
Poly(ADP-ribose) polymerase-1 regulates the stability of the wild-type p53 protein.Cell Mol Biol Lett. 2001;6(2):117-40. Cell Mol Biol Lett. 2001. PMID: 11544635
-
DNA repair factors and telomere-chromosome integrity in mammalian cells.Cytogenet Genome Res. 2004;104(1-4):116-22. doi: 10.1159/000077475. Cytogenet Genome Res. 2004. PMID: 15162024 Review.
-
[Analysis of the role of DNA repair, regulation of cell cycle and apoptosis in the radiation-induced adaptive response of mammalian cells].Radiats Biol Radioecol. 2003 Jan-Feb;43(1):19-28. Radiats Biol Radioecol. 2003. PMID: 12677654 Review. Russian.
Cited by
-
Global analysis of transcriptional regulation by poly(ADP-ribose) polymerase-1 and poly(ADP-ribose) glycohydrolase in MCF-7 human breast cancer cells.J Biol Chem. 2009 Dec 4;284(49):33926-38. doi: 10.1074/jbc.M109.023879. Epub 2009 Oct 7. J Biol Chem. 2009. PMID: 19812418 Free PMC article.
-
Reviewing the characteristics of BRCA and PALB2-related cancers in the precision medicine era.Genet Mol Biol. 2019;42(1 suppl 1):215-231. doi: 10.1590/1678-4685-GMB-2018-0104. Epub 2019 Apr 29. Genet Mol Biol. 2019. PMID: 31067289 Free PMC article.
-
PARP-1 Val762Ala polymorphism is associated with reduced risk of non-Hodgkin lymphoma in Korean males.BMC Med Genet. 2010 Mar 3;11:38. doi: 10.1186/1471-2350-11-38. BMC Med Genet. 2010. PMID: 20196871 Free PMC article.
-
PARP-1 and Ku compete for repair of DNA double strand breaks by distinct NHEJ pathways.Nucleic Acids Res. 2006;34(21):6170-82. doi: 10.1093/nar/gkl840. Epub 2006 Nov 6. Nucleic Acids Res. 2006. PMID: 17088286 Free PMC article.
-
New facets in the regulation of gene expression by ADP-ribosylation and poly(ADP-ribose) polymerases.Chem Rev. 2015 Mar 25;115(6):2453-81. doi: 10.1021/cr5004248. Epub 2015 Jan 9. Chem Rev. 2015. PMID: 25575290 Free PMC article. Review. No abstract available.
References
-
- Ambs S. et al. (1998a) p53 and vascular endothelial growth factor regulate tumor growth of NOS2-expressing human carcinoma cells. Nature Med., 4, 1371–1376. - PubMed
-
- Bauer P.I. et al. (1996) Modification of growth related enzymatic pathways and apparent loss of tumorigenicity of a ras-transformed bovine endothelial cell line by treatment with 5-iodo-6-amino-1,2-benzopyrone (INH2BP). Int. J. Oncol., 8, 239–252. - PubMed
-
- Berger N.A. (1985) Poly(ADP-ribose) in the cellular response to DNA damage. Radiat. Res., 101, 4–15. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
