

Functional characterization of the C-terminus of the human ether-à-go-go-related gene K(+) channel (HERG)
- PMID: 11432987
- PMCID: PMC2278693
- DOI: 10.1111/j.1469-7793.2001.t01-3-00001.x
Functional characterization of the C-terminus of the human ether-à-go-go-related gene K(+) channel (HERG)
Abstract
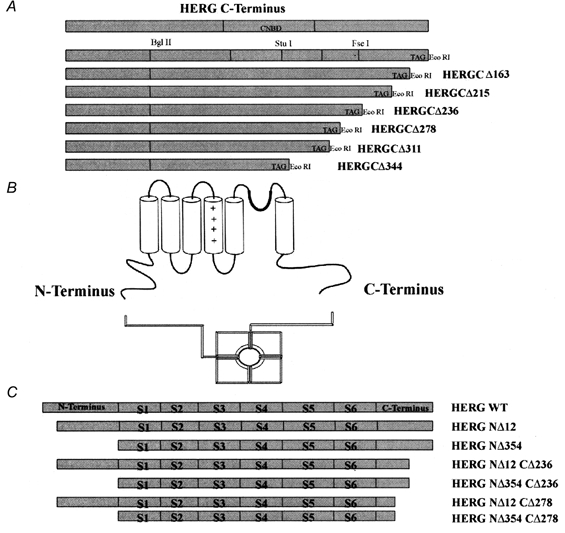
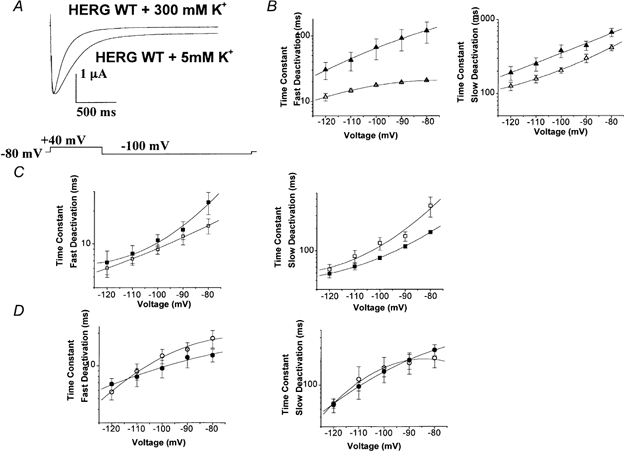
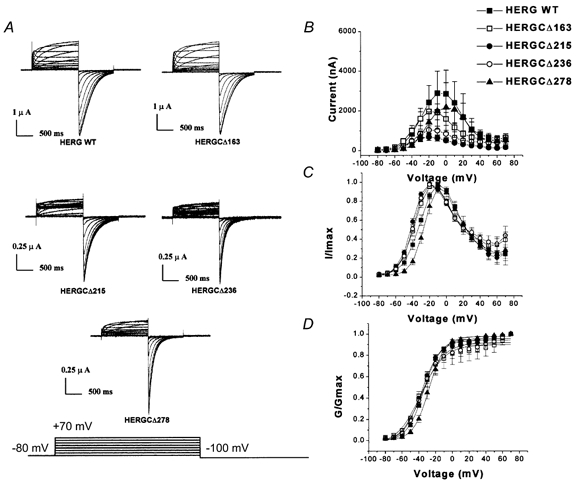
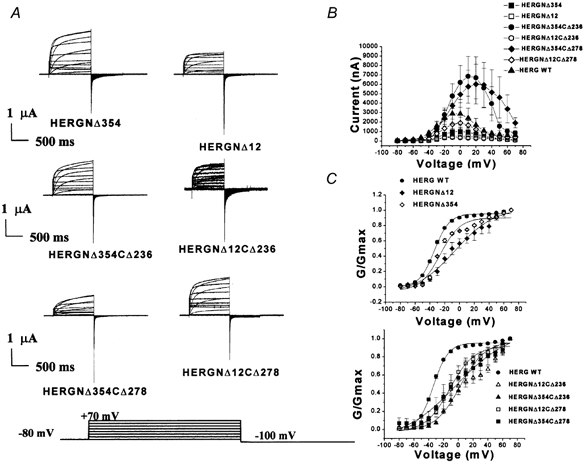
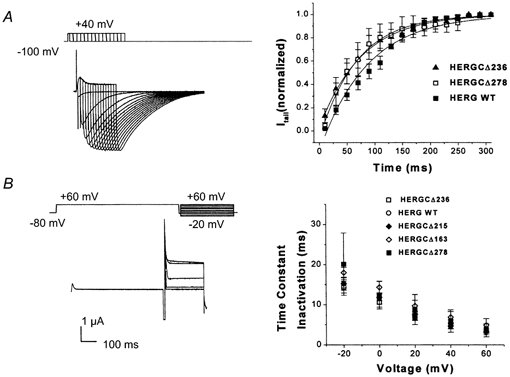
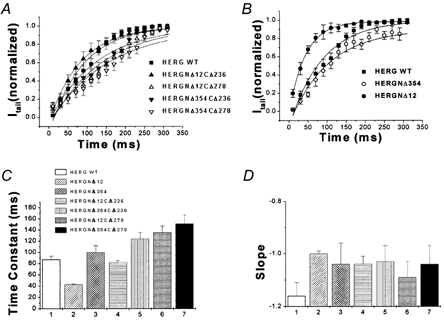
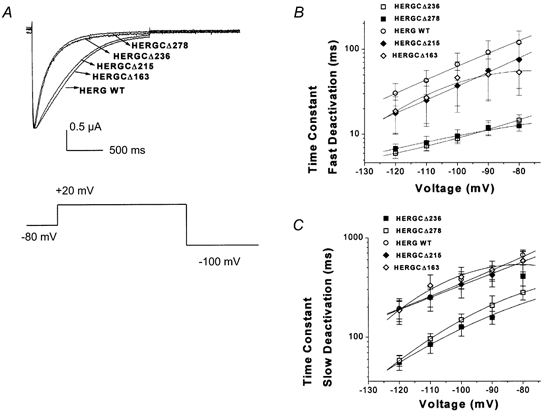
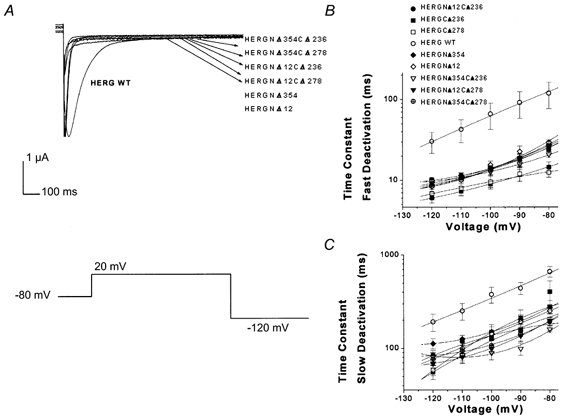
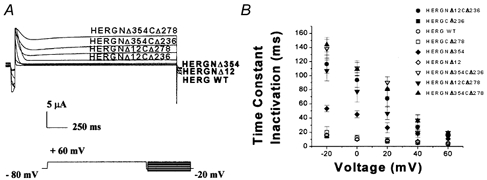
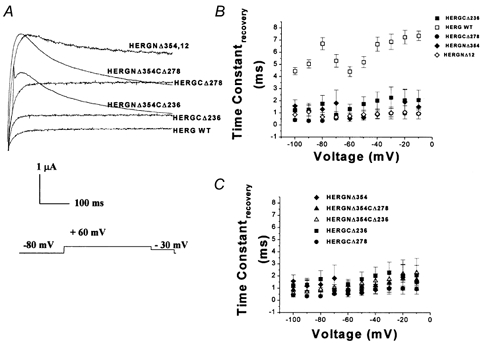
1. In the present study the functional role of the C-terminus of the human ether-à-go-go-related gene K(+) channel HERG was investigated using a series of C-terminal deletion constructs expressed in Xenopus oocytes. 2. Constructs with deletions of 311 or more amino acid residues failed to form functional channels. Truncation by 215 amino acid residues or fewer had no discernable effects on channel activity. Truncation by 236 or 278 amino acid residues accelerated deactivation, and caused a faster recovery from inactivation. 3. In high extracellular K(+), channel deactivation of HERG results from the binding of the N-terminus to a site within the pore. This slows channel deactivation by a knock-off mechanism. Here, it was shown that C-terminal deletions also abolished this effect of high extracellular K(+). Mutants containing deletions in both the N- and C-termini deactivated with rates similar to those observed in individual deletion mutants. 4. In contrast, experiments with double-deletion constructs showed additive effects of the N- and C-termini on the voltage dependence of activation, and on the kinetics of inactivation and recovery from inactivation. The reduction of inactivation in these mutants contributed to an increase in peak current amplitude. 5. These results indicate that residues within the C-terminus of HERG play a role in channel expression as well as in most aspects of channel gating. The regulation of channel deactivation is likely to be mediated by an interaction with the N-terminus, but the regulation of the voltage dependence of activation, and of rate processes associated with inactivation, does not require the N-terminus.
Figures
References
-
- Abbott GW, Sesti F, Splawski I, Buck ME, Lehmann MH, Timothy KW, Keating MT, Goldstein SA. MiRP1 forms IKr potassium channels with HERG and is associated with cardiac arrhythmia. Cell. 1999;97:175–187. - PubMed
-
- Arnold DB, Clapham DE. Molecular determinants for subcellular localization of PSD-95 with an interacting K+ channel. Neuron. 1999;23:149–157. - PubMed
-
- Bentley GN, Brooks MA, O'Neill CA, Findlay JB. Determinants of potassium channel assembly localised within the cytoplasmic C-terminal domain of Kv2. 1. Biochimica et Biophysica Acta. 1999;1418:176–184. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources