

African swine fever virus structural protein pE120R is essential for virus transport from assembly sites to plasma membrane but not for infectivity
- PMID: 11435554
- PMCID: PMC114402
- DOI: 10.1128/JVI.75.15.6758-6768.2001
African swine fever virus structural protein pE120R is essential for virus transport from assembly sites to plasma membrane but not for infectivity
Abstract
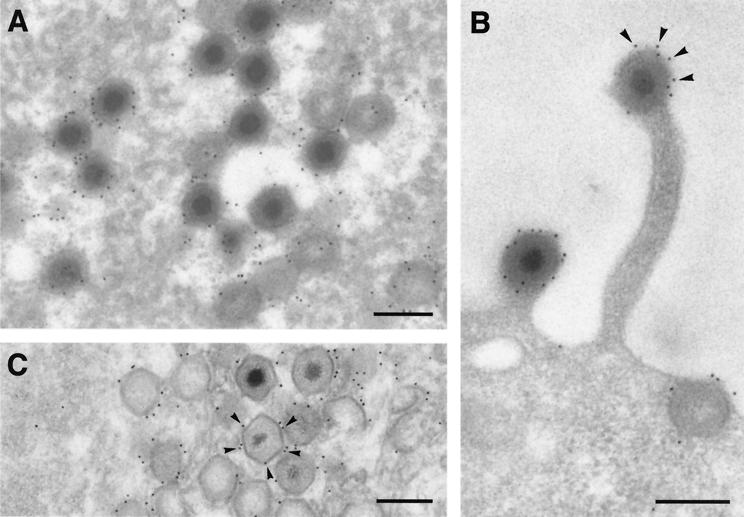
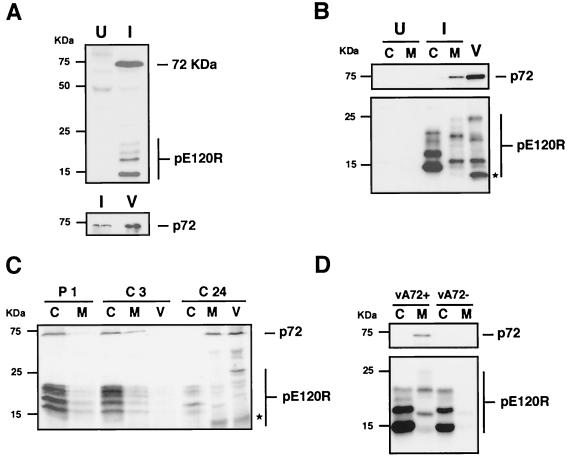
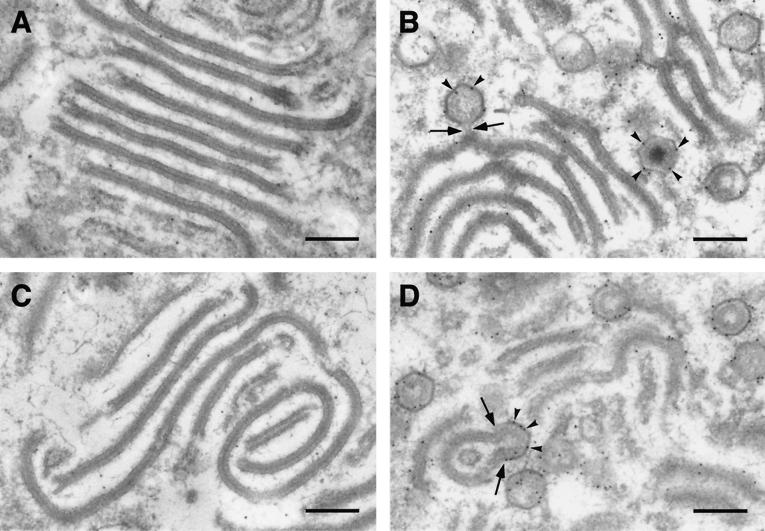
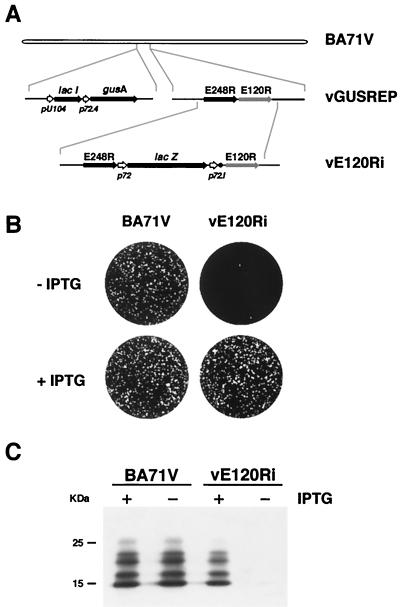
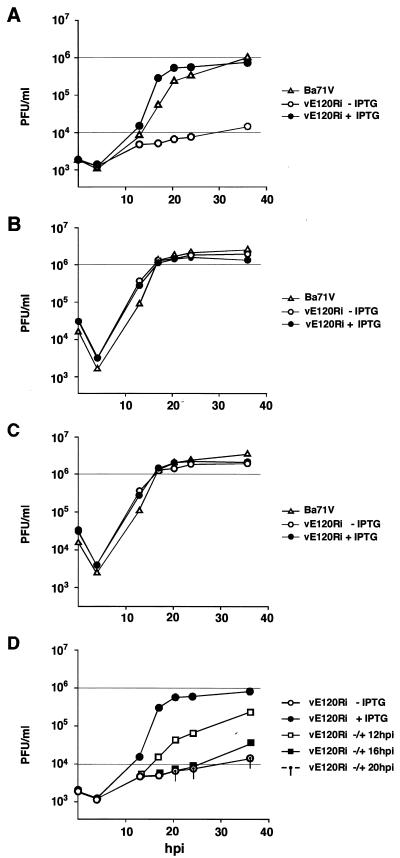
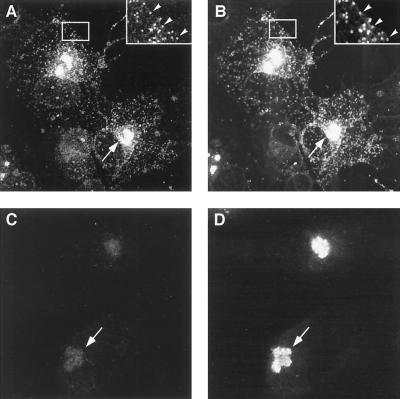
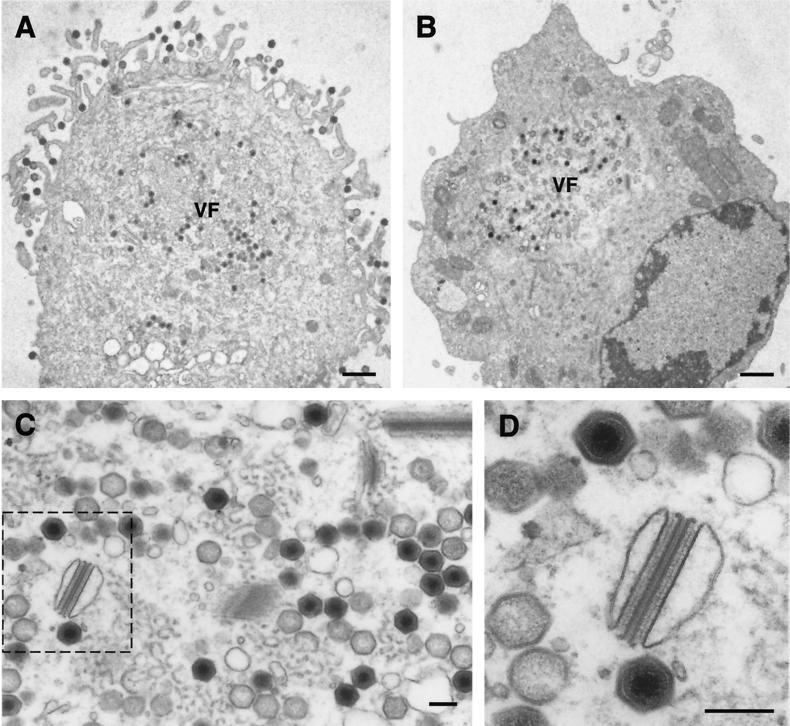
This report examines the role of African swine fever virus (ASFV) structural protein pE120R in virus replication. Immunoelectron microscopy revealed that protein pE120R localizes at the surface of the intracellular virions. Consistent with this, coimmunoprecipitation assays showed that protein pE120R binds to the major capsid protein p72. Moreover, it was found that, in cells infected with an ASFV recombinant that inducibly expresses protein p72, the incorporation of pE120R into the virus particle is dependent on p72 expression. Protein pE120R was also studied using an ASFV recombinant in which E120R gene expression is regulated by the Escherichia coli lac repressor-operator system. In the absence of inducer, pE120R expression was reduced about 100-fold compared to that obtained with the parental virus or the recombinant virus grown under permissive conditions. One-step virus growth curves showed that, under conditions that repress pE120R expression, the titer of intracellular progeny was similar to the total virus yield obtained under permissive conditions, whereas the extracellular virus yield was about 100-fold lower than in control infections. Immunofluorescence and electron microscopy demonstrated that, under restrictive conditions, intracellular mature virions are properly assembled but remain confined to the replication areas. Altogether, these results indicate that pE120R is necessary for virus dissemination but not for virus infectivity. The data also suggest that protein pE120R might be involved in the microtubule-mediated transport of ASFV particles from the viral factories to the plasma membrane.
Figures
References
-
- Alves de Matos A P, Carvalho Z G. African swine fever virus interaction with microtubules. Biol Cell. 1993;78:229–234. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
