

BeF(3)(-) acts as a phosphate analog in proteins phosphorylated on aspartate: structure of a BeF(3)(-) complex with phosphoserine phosphatase
- PMID: 11438683
- PMCID: PMC37469
- DOI: 10.1073/pnas.131213698
BeF(3)(-) acts as a phosphate analog in proteins phosphorylated on aspartate: structure of a BeF(3)(-) complex with phosphoserine phosphatase
Abstract
Protein phosphoaspartate bonds play a variety of roles. In response regulator proteins of two-component signal transduction systems, phosphorylation of an aspartate residue is coupled to a change from an inactive to an active conformation. In phosphatases and mutases of the haloacid dehalogenase (HAD) superfamily, phosphoaspartate serves as an intermediate in phosphotransfer reactions, and in P-type ATPases, also members of the HAD family, it serves in the conversion of chemical energy to ion gradients. In each case, lability of the phosphoaspartate linkage has hampered a detailed study of the phosphorylated form. For response regulators, this difficulty was recently overcome with a phosphate analog, BeF(3)(-), which yields persistent complexes with the active site aspartate of their receiver domains. We now extend the application of this analog to a HAD superfamily member by solving at 1.5-A resolution the x-ray crystal structure of the complex of BeF(3)(-) with phosphoserine phosphatase (PSP) from Methanococcus jannaschii. The structure is comparable to that of a phosphoenzyme intermediate: BeF(3)(-) is bound to Asp-11 with the tetrahedral geometry of a phosphoryl group, is coordinated to Mg(2+), and is bound to residues surrounding the active site that are conserved in the HAD superfamily. Comparison of the active sites of BeF(3)(-) x PSP and BeF(3)(-) x CeY, a receiver domain/response regulator, reveals striking similarities that provide insights into the function not only of PSP but also of P-type ATPases. Our results indicate that use of BeF(3)(-) for structural studies of proteins that form phosphoaspartate linkages will extend well beyond response regulators.
Figures
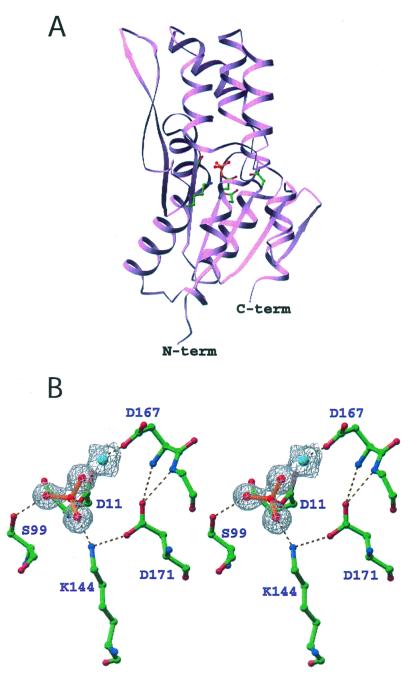
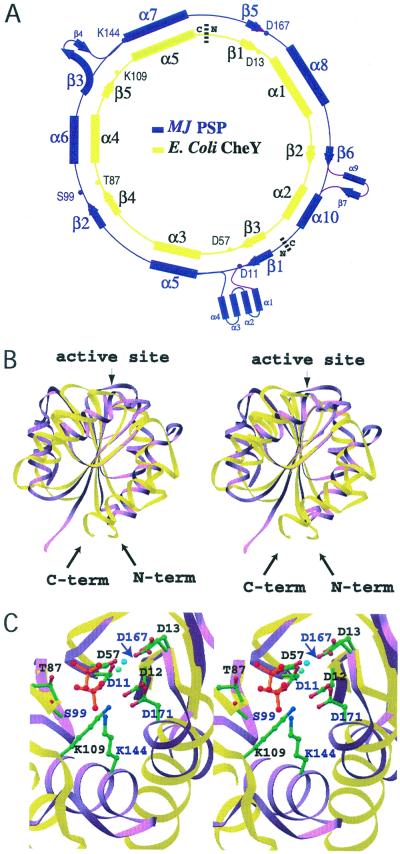
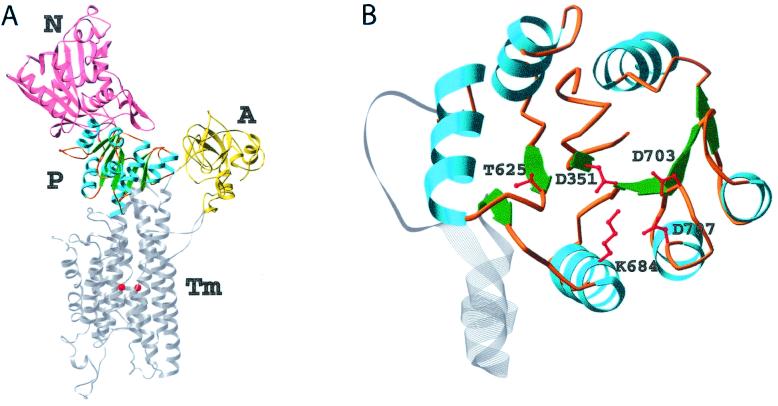
Comment in
-
Probing the mechanism of enzymatic phosphoryl transfer with a chemical trick.Proc Natl Acad Sci U S A. 2001 Jul 17;98(15):8170-1. doi: 10.1073/pnas.161273998. Proc Natl Acad Sci U S A. 2001. PMID: 11459948 Free PMC article. Review. No abstract available.
References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
