

Review
doi: 10.1093/emboj/20.14.3609.
Apical-basal pattern formation in Arabidopsis embryogenesis
Affiliations
- PMID: 11447101
- PMCID: PMC125542
- DOI: 10.1093/emboj/20.14.3609
Item in Clipboard
Review
Apical-basal pattern formation in Arabidopsis embryogenesis
EMBO J.
.
No abstract available
Figures
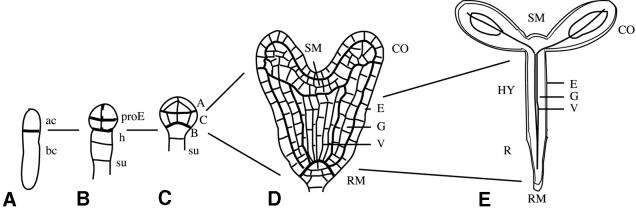
Fig. 1. Development of the apical–basal pattern during Arabidopsis embryogenesis. (A) One-cell stage. The zygote has divided asymmetrically into an apical (ac) and a basal (bc) daughter cell. (B) Octant stage. The proembryo (proE) derived from the apical cell consists of two tiers each of four cells. The basal cell has produced a file of cells, including the hypophysis (h) and the suspensor (su). (C) Dermatogen stage. Three embryo regions are indicated: A, apical; C, central; B, basal. (D) Heart stage. The basic body organization is in place. SM, shoot meristem; CO, cotyledon primordia; RM, root meristem; E, epidermis; G, ground tissue; V, vascular primordium. (E) Seedling. HY, hypocotyl; R, root. Lines indicate the origin of seedling structures from early embryo regions.
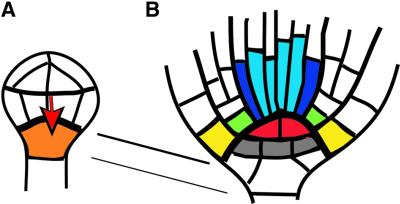
Fig. 2. Origin of the primary root meristem during embryogenesis. (A) Dermatogen stage. A signal from the proembryo (red arrow) is presumed to induce the cell fate of the hypophysis (orange). (B) Bottom end of heart-stage embryo. The quiescent centre (red) presumably induces stem-cell fate of surrounding cells. The upper tier of stem cells (colour coded) will produce root tissue cells; the lower tier of stem cells (grey) will form central root cap cells. The connecting lines indicate descent of the quiescent centre and the lower tier stem cells from the hypophysis.
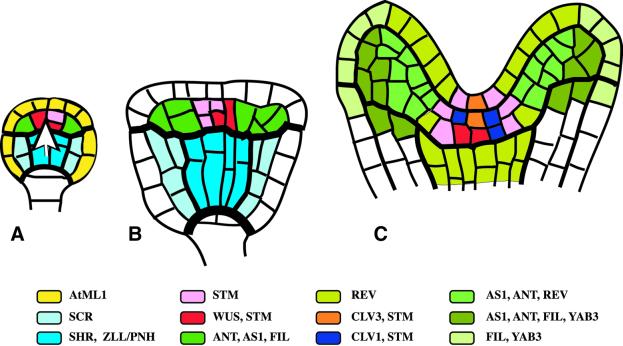
Fig. 3. Origin of the primary shoot meristem and the shoot apical organization. Approximate domains of gene expression are colour coded. (A) Globular embryo. The epidermis-specific AtML1 gene is expressed in both the central and the apical region (yellow; not shown in B and C). The white arrow indicates a cue from the central region in positioning of the shoot meristem primordium. (B) Transition-stage embryo displaying essentially the same expression pattern as the globular embryo. (C) Top end of heart-stage embryo. Expression domains of radial patterning genes, SCR and SHR, and of ZLL/PNH are not shown. In the shoot meristem primordium, the expression domains of CLV3 and CLV1 overlap. Note adaxial (REV) and abaxial (FIL, YAB3) gene expression domains in the cotyledon primordia. REV is also expressed in the vascular primordium.
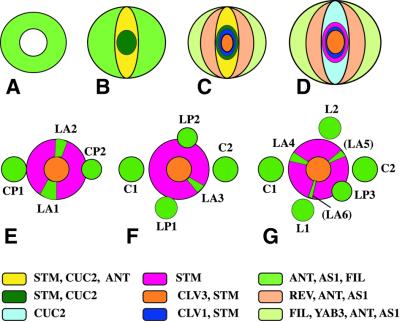
Fig. 4. Origin of phyllotaxis in the shoot apex. (A–D) Schematic cross-section through the apical region of the embryo. (A) Globular stage, with ring-shaped expression domain of ANT, AS1, FIL in the periphery. (B) Transition stage, showing expression stripe of STM and CUC2 across the apex. (C) Heart stage. CLV1 and CLV3 are expressed in the centre, and the periphery is subdivided into adaxial (REV) and abaxial (FIL) expression domains. (D) Walking-stick stage. CUC2 and STM expression domains become mutually exclusive. (E–G) Gradual transition from embryonic to post-embryonic phyllotaxis. The angles separating successive primordia are approximately: 180° (C1, C2), 100° (C2, L1), 175° (L1, L2), 110° (L2, L3), 160° (L3, L4), 130° (L4, L5) and 140° (L5, L6). Leaf primordium initiation sites (green sectors) within the peripheral zone of the shoot meristem are highlighted by expression of PIN1, AN, AS1, REV and FIL and by absence of STM expression (see text). C, cotyledon; CP, cotyledon primordium; L, leaf; LP, leaf primordium; LA, leaf anlage (founder cells at initiation site). Young primordia bordering on the peripheral zone have inhibitory effects on the initiation of the next primordium.
References
-
- Aida M., Ishida,T. and Tasaka M. (1999) Shoot apical meristem and cotyledon formation during Arabidopsis embryogenesis: interaction among the CUP-SHAPED COTYLEDON and SHOOT MERI STEMLESS genes. Development, 126, 1563–1570. - PubMed
-
- Barton M.K. and Poethig,R.S. (1993) Formation of the shoot apical meristem in Arabidopsis thaliana: an analysis of development in the wild type and in the shoot meristemless mutant. Development, 119, 823–831.
-
- Berleth T. and Jürgens,G. (1993) The role of the monopteros gene in organising the basal body region of the Arabidopsis embryo. Development, 118, 575–587.
-
- Brand U., Fletcher,J.C., Hobe,M., Meyerowitz,E.M. and Simon,R. (2000) Dependence of stem cell fate in Arabidopsis on a feedback loop regulated by CLV3 activity. Science, 289, 817–819. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
