

Adenylyl cyclase Rv1625c of Mycobacterium tuberculosis: a progenitor of mammalian adenylyl cyclases
- PMID: 11447108
- PMCID: PMC125536
- DOI: 10.1093/emboj/20.14.3667
Adenylyl cyclase Rv1625c of Mycobacterium tuberculosis: a progenitor of mammalian adenylyl cyclases
Abstract
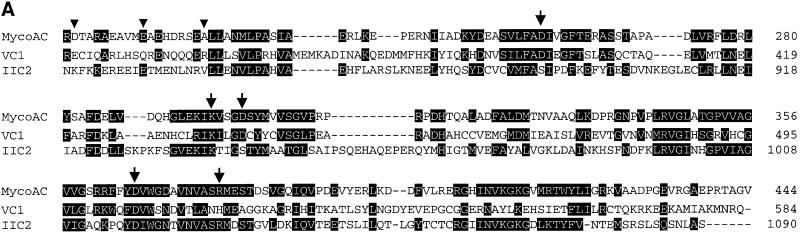
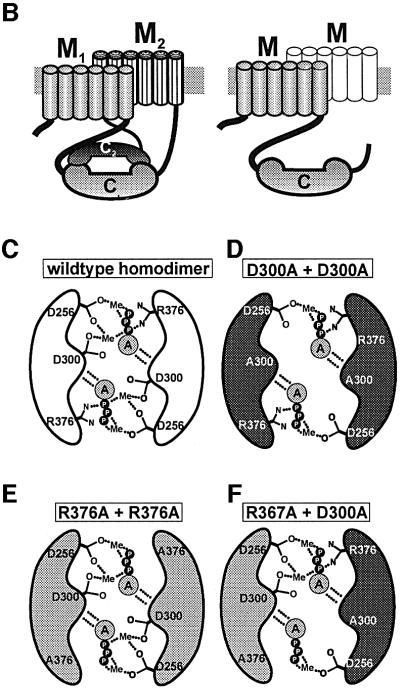
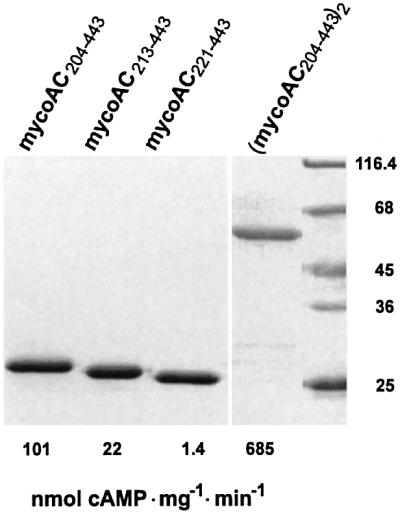
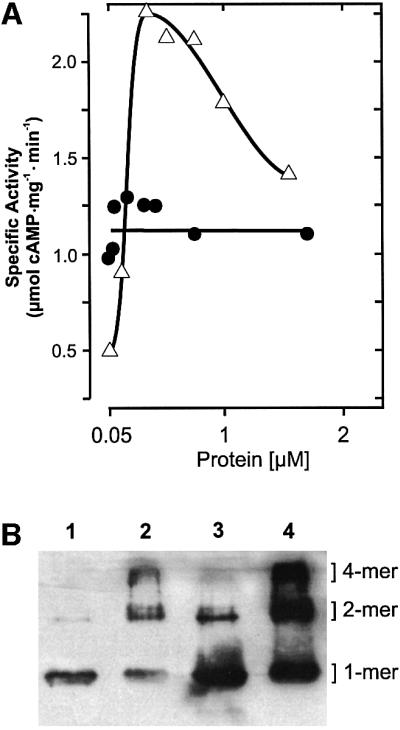
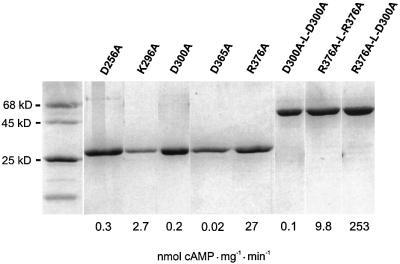
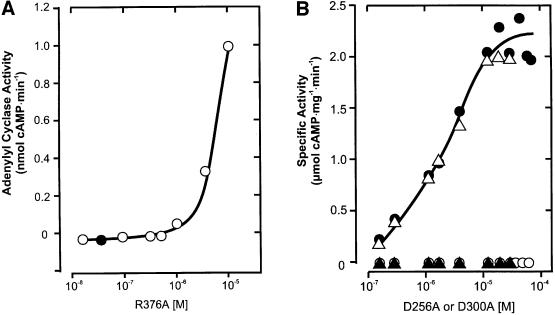
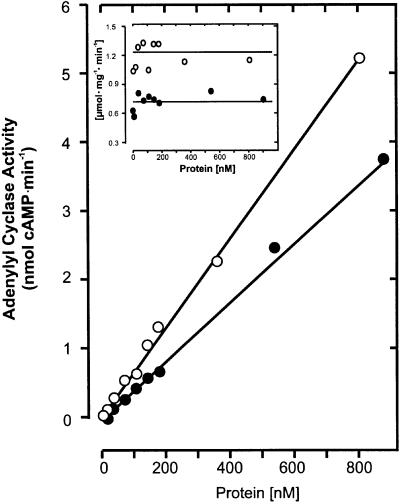
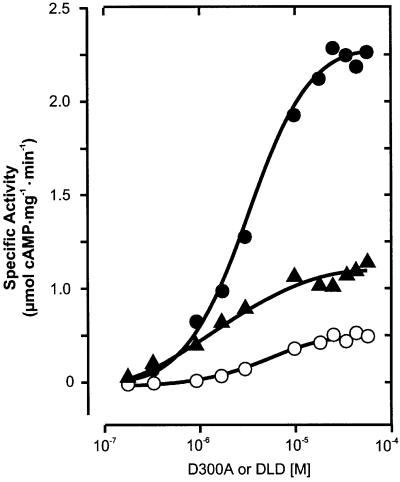
The gene Rv1625c from Mycobacterium tuberculosis encodes a membrane-anchored adenylyl cyclase corresponding to exactly one-half of a mammalian adenylyl cyclase. An engineered, soluble form of Rv1625c was expressed in Escherichia coli. It formed a homodimeric cyclase with two catalytic centers. Amino acid mutations predicted to affect catalysis resulted in inactive monomers. A single catalytic center with wild-type activity could be reconstituted from mutated monomers in stringent analogy to the mammalian heterodimeric cyclase structure. The proposed existence of supramolecular adenylyl cyclase complexes was established by reconstitution from peptide-linked, mutation-inactivated homodimers resulting in pseudo-trimeric and -tetrameric complexes. The mycobacterial holoenzyme was expressed successfully in E.coli and mammalian HEK293 cells, i.e. its membrane targeting sequence was compatible with the bacterial and eukaryotic machinery for processing and membrane insertion. The membrane-anchored mycobacterial cyclase expressed in E.coli was purified to homogeneity as a first step toward the complete structural elucidation of this important protein. As the closest progenitor of the mammalian adenylyl cyclase family to date, the mycobacterial cyclase probably was spread by horizontal gene transfer.
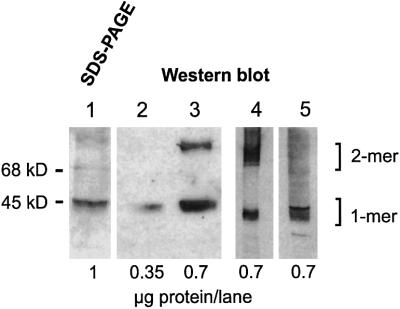
Figures
References
-
- Baltimore D. (2001) Our genome unveiled. Nature, 409, 814–816. - PubMed
-
- Bhatnagar N.B., Bhatnagar,R. and Venkitasubramanian,T.A. (1984) Characterization and metabolism of cyclic guanosine 3′,5′-monophosphate in Mycobacterium smegmatis. Biochem. Biophys. Res. Commun., 121, 634–640. - PubMed
-
- Chang G., Spencer,R.H., Lee,A.T., Barclay,M.T. and Rees,D.C. (1998) Structure of the MscL homolog from Mycobacterium tuberculosis: a gated mechanosensitive ion channel. Science, 282, 2220–2226. - PubMed
-
- Chen G.Q., Cui,C., Mayer,M.L. and Gouaux,E. (1999) Functional characterization of a potassium-selective prokaryotic glutamate receptor. Nature, 402, 817–821. - PubMed
-
- Chen J. et al. (1995) A region of adenylyl cyclase 2 critical for regulation by G protein βγ subunits. Science, 268, 1166–1169. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
