

Escherichia coli enterotoxin B subunit triggers apoptosis of CD8(+) T cells by activating transcription factor c-myc
- PMID: 11447169
- PMCID: PMC98583
- DOI: 10.1128/IAI.69.8.4923-4930.2001
Escherichia coli enterotoxin B subunit triggers apoptosis of CD8(+) T cells by activating transcription factor c-myc
Abstract
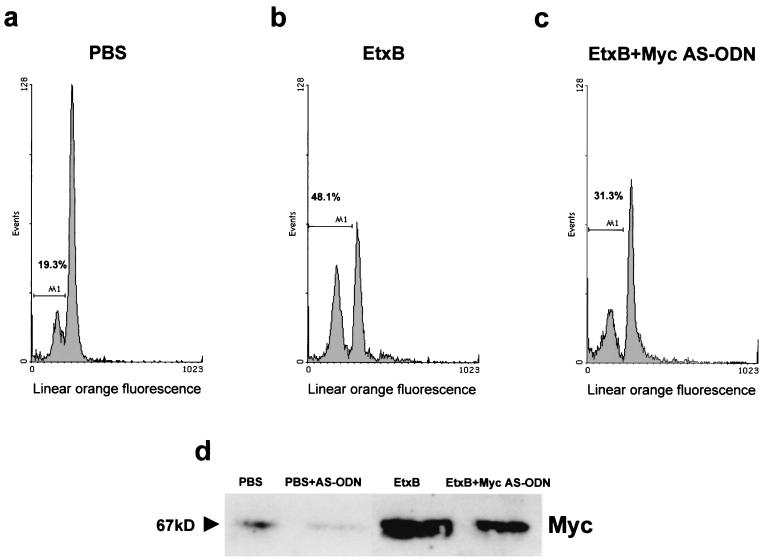
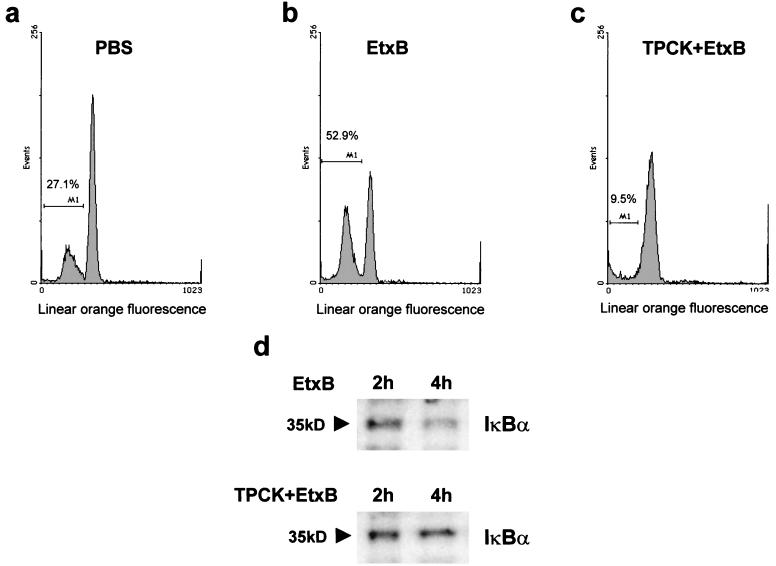
Heat-labile enterotoxin from enterotoxinogenic Escherichia coli is not only an important cause of diarrhea in humans and domestic animals but also possesses potent immunomodulatory properties. Recently, the nontoxic, receptor-binding B subunit of heat-labile enterotoxin (EtxB) was found to induce the selective death of CD8(+) T cells, suggesting that EtxB may trigger activation of proapoptotic signaling pathways. Here we show that EtxB treatment of CD8(+) T cells but not of CD4(+) T cells triggers the specific up-regulation of the transcription factor c-myc, implicated in the control of cell proliferation, differentiation, and death. A concomitant elevation in Myc protein levels was also evident, with peak expression occurring 4 h posttreatment. Preincubation with c-myc antisense oligodeoxynucleotides demonstrated that Myc expression was necessary for EtxB-mediated apoptosis. Myc activation was also associated with an increase of IkappaBalpha turnover, suggesting that elevated Myc expression may be dependent on NF-kappaB. When CD8(+) T cells were pretreated with inhibitors of IkappaBalpha turnover and NF-kappaB translocation, this resulted in a marked reduction in both EtxB-induced apoptosis and Myc expression. Further, a non-receptor-binding mutant of EtxB, EtxB(G33D), was shown to lack the capacity to activate Myc transcription. These findings provide further evidence that EtxB is a signaling molecule that triggers activation of transcription factors involved in cell survival.
Figures
Similar articles
-
The B subunit of Escherichia coli heat-labile enterotoxin induces both caspase-dependent and -independent cell death pathways in CD8+ T cells.Infect Immun. 2004 Oct;72(10):5850-7. doi: 10.1128/IAI.72.10.5850-5857.2004. Infect Immun. 2004. PMID: 15385486 Free PMC article.
-
Mutant Escherichia coli heat-labile toxin B subunit that separates toxoid-mediated signaling and immunomodulatory action from trafficking and delivery functions.Infect Immun. 2003 Mar;71(3):1527-37. doi: 10.1128/IAI.71.3.1527-1537.2003. Infect Immun. 2003. PMID: 12595472 Free PMC article.
-
CD8+ T cell apoptosis induced by Escherichia coli heat-labile enterotoxin B subunit occurs via a novel pathway involving NF-kappaB-dependent caspase activation.Eur J Immunol. 2002 Jun;32(6):1737-47. doi: 10.1002/1521-4141(200206)32:6<1737::AID-IMMU1737>3.0.CO;2-J. Eur J Immunol. 2002. PMID: 12115657
-
Cross-linking of cell surface ganglioside GM1 induces the selective apoptosis of mature CD8+ T lymphocytes.Int Immunol. 1996 May;8(5):731-6. doi: 10.1093/intimm/8.5.731. Int Immunol. 1996. PMID: 8671661 Review.
-
Immune modulation by the cholera-like enterotoxin B-subunits: from adjuvant to immunotherapeutic.Int J Med Microbiol. 2000 Oct;290(4-5):447-53. doi: 10.1016/S1438-4221(00)80062-4. Int J Med Microbiol. 2000. PMID: 11111925 Review.
Cited by
-
Gangliosides in T cell development and function of mice.Glycoconj J. 2022 Apr;39(2):229-238. doi: 10.1007/s10719-021-10037-5. Epub 2022 Feb 14. Glycoconj J. 2022. PMID: 35156158 Review.
-
The B subunit of Escherichia coli heat-labile enterotoxin induces both caspase-dependent and -independent cell death pathways in CD8+ T cells.Infect Immun. 2004 Oct;72(10):5850-7. doi: 10.1128/IAI.72.10.5850-5857.2004. Infect Immun. 2004. PMID: 15385486 Free PMC article.
-
Mutant Escherichia coli heat-labile toxin B subunit that separates toxoid-mediated signaling and immunomodulatory action from trafficking and delivery functions.Infect Immun. 2003 Mar;71(3):1527-37. doi: 10.1128/IAI.71.3.1527-1537.2003. Infect Immun. 2003. PMID: 12595472 Free PMC article.
-
The B subunit of Escherichia coli heat-labile enterotoxin enhances CD8+ cytotoxic-T-lymphocyte killing of Epstein-Barr virus-infected cell lines.J Virol. 2003 Apr;77(7):4298-305. doi: 10.1128/jvi.77.7.4298-4305.2003. J Virol. 2003. PMID: 12634387 Free PMC article.
-
Different types of cell death induced by enterotoxins.Toxins (Basel). 2010 Aug;2(8):2158-76. doi: 10.3390/toxins2082158. Epub 2010 Aug 11. Toxins (Basel). 2010. PMID: 22069678 Free PMC article. Review.
References
-
- Amati B, Brooks M W, Levy N, Littlewood T D, Evan G I, Land H. Oncogenic activity of the c-Myc protein requires dimerization with Max. Cell. 1993;72:233–245. - PubMed
-
- Amati B, Land H. Myc-Max-Mad: a transcription factor network controlling cell cycle progression, differentiation and death. Curr Opin Genet Dev. 1994;4:102–108. - PubMed
-
- Ayer D E, Eisenman R N. A switch from Myc:Max to Mad:Max heterocomplexes accompanies monocyte/macrophage differentiation. Genes Dev. 1993;7:2110–2119. - PubMed
-
- Ayer D E, Kretzner L, Eisenman R N. Mad: a heterodimeric partner for Max that antagonizes Myc transcriptional activity. Cell. 1993;72:211–222. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
