

Mechanism of DNA polymerase II-mediated frameshift mutagenesis
- PMID: 11447256
- PMCID: PMC37476
- DOI: 10.1073/pnas.141113398
Mechanism of DNA polymerase II-mediated frameshift mutagenesis
Abstract
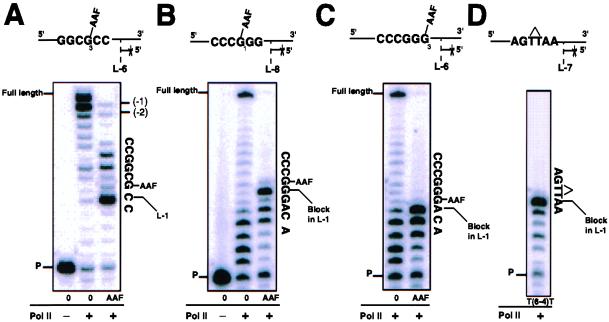
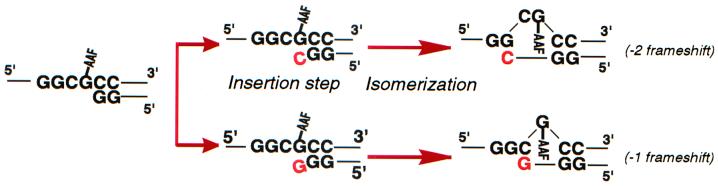
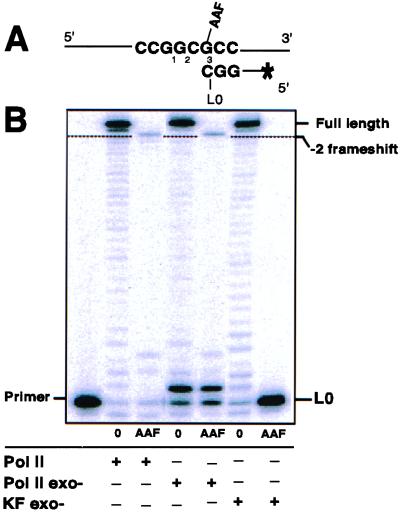
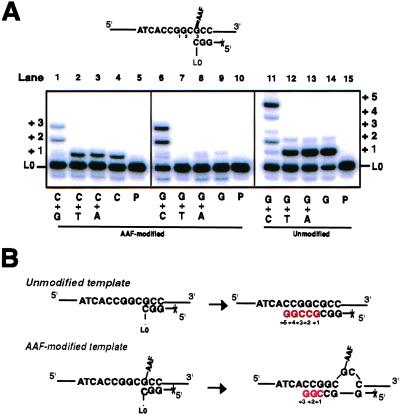
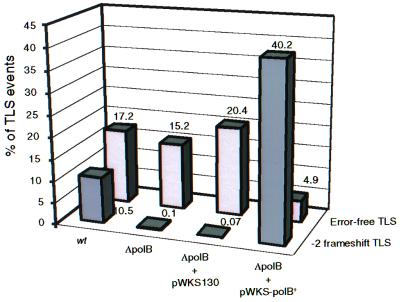
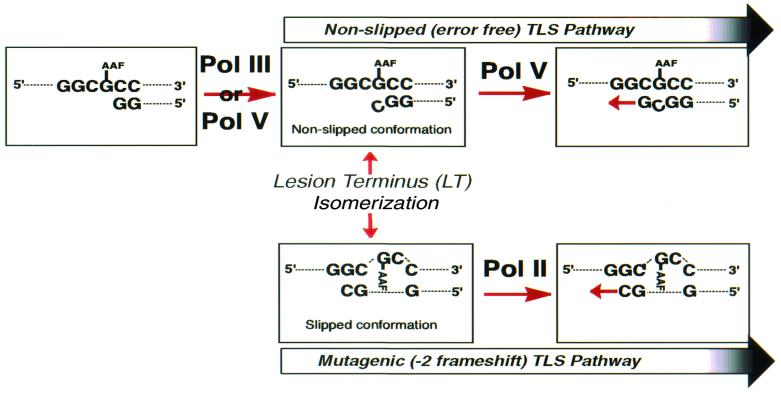
Escherichia coli possesses three SOS-inducible DNA polymerases (Pol II, IV, and V) that were recently found to participate in translesion synthesis and mutagenesis. Involvement of these polymerases appears to depend on the nature of the lesion and its local sequence context, as illustrated by the bypass of a single N-2-acetylaminofluorene adduct within the NarI mutation hot spot. Indeed, error-free bypass requires Pol V (umuDC), whereas mutagenic (-2 frameshift) bypass depends on Pol II (polB). In this paper, we show that purified DNA Pol II is able in vitro to generate the -2 frameshift bypass product observed in vivo at the NarI sites. Although the Delta polB strain is completely defective in this mutation pathway, introduction of the polB gene on a low copy number plasmid restores the -2 frameshift pathway. In fact, modification of the relative copy number of polB versus umuDC genes results in a corresponding modification in the use of the frameshift versus error-free translesion pathways, suggesting a direct competition between Pol II and V for the bypass of the same lesion. Whether such a polymerase competition model for translesion synthesis will prove to be generally applicable remains to be confirmed.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
