

Contact interactions between epitheliocytes and fibroblasts: formation of heterotypic cadherin-containing adhesion sites is accompanied by local cytoskeletal reorganization
- PMID: 11447275
- PMCID: PMC37487
- DOI: 10.1073/pnas.151247698
Contact interactions between epitheliocytes and fibroblasts: formation of heterotypic cadherin-containing adhesion sites is accompanied by local cytoskeletal reorganization
Abstract
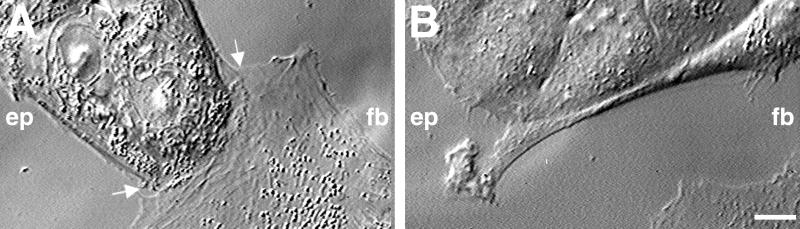
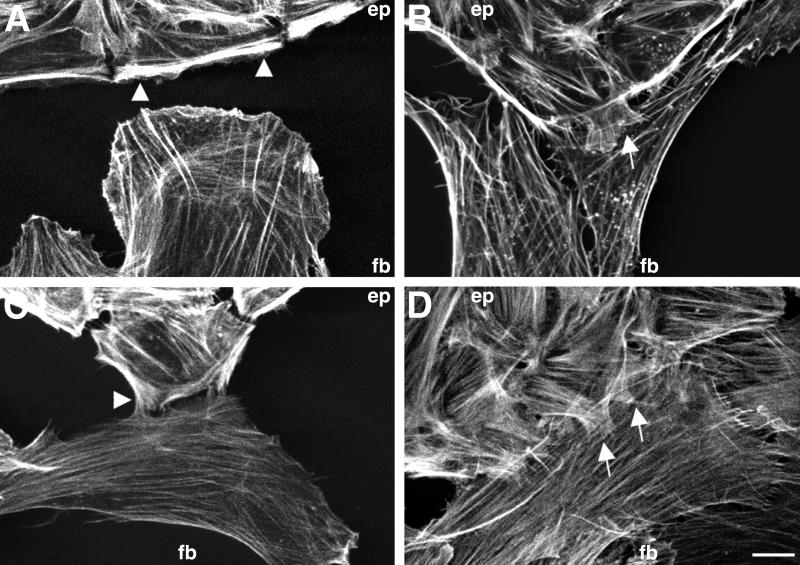
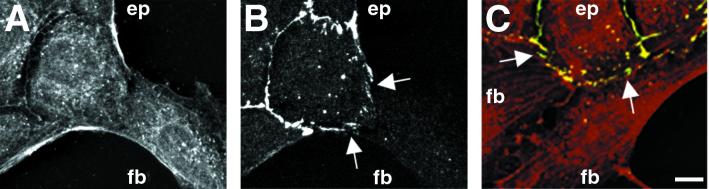
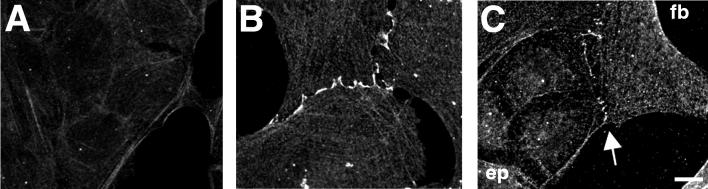
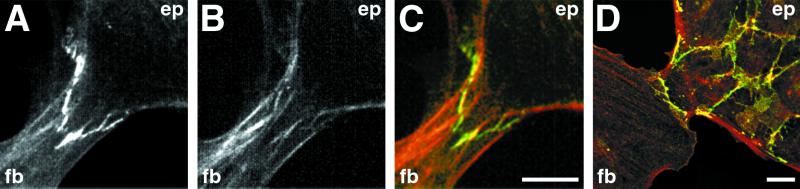
Contact interactions between different cell types play a number of important roles in development, for example in cell sorting, tissue organization, and ordered migration of cells. The nature of such heterocellular interactions, in contrast to interactions between cells of the same type, remains largely unknown. In this report, we present experimental data examining the dynamics of heterocellular interactions between epitheliocytes and fibroblasts, which express different cadherin cell adhesion molecules and possess different actin cytoskeletal organizations. Our analysis revealed two striking features of heterocellular contact. First, the active free edge of an epitheliocyte reorganizes its actin cytoskeleton after making contact with a fibroblast. Upon contact with the leading edge of a fibroblast, epitheliocytes disassemble their marginal bundle of actin filaments and reassemble actin filaments into a geometric organization more typical of a fibroblast lamella. Second, epitheliocytes and fibroblasts form cell--cell adhesion structures that have an irregular organization and are associated with components of cell adhesion complexes. The structural organization of these adhesions is more closely related to the type of contacts formed between fibroblasts rather than to those between epitheliocytes. Heterotypic epithelio-fibroblastic contacts, like homotypic contacts between fibroblasts, are transient and do not lead to formation of stable contact interactions. We suggest that heterocellular contact interactions in culture may be regarded as models of how tissue systems consisting of epithelia and mesenchyme interact and become organized in vivo.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
