

The infected cell protein 0 of herpes simplex virus 1 dynamically interacts with proteasomes, binds and activates the cdc34 E2 ubiquitin-conjugating enzyme, and possesses in vitro E3 ubiquitin ligase activity
- PMID: 11447293
- PMCID: PMC37518
- DOI: 10.1073/pnas.161283098
The infected cell protein 0 of herpes simplex virus 1 dynamically interacts with proteasomes, binds and activates the cdc34 E2 ubiquitin-conjugating enzyme, and possesses in vitro E3 ubiquitin ligase activity
Abstract
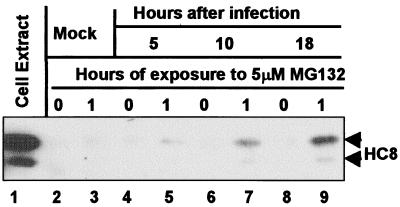
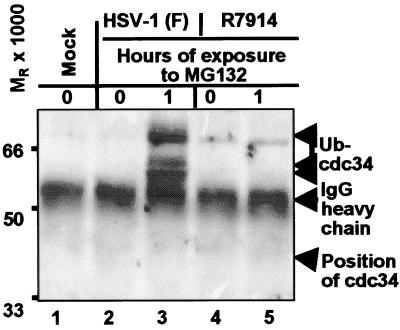
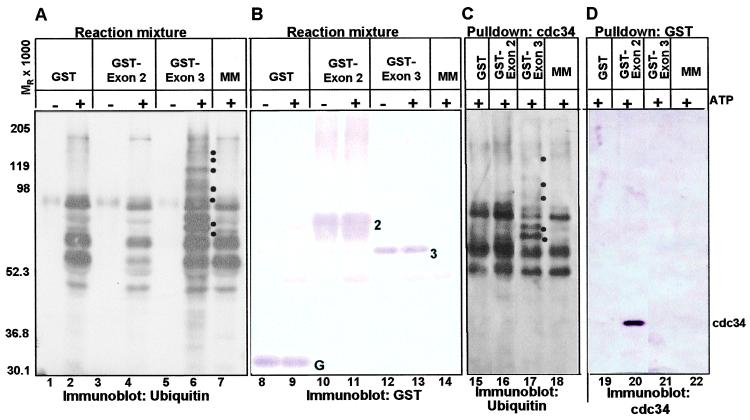
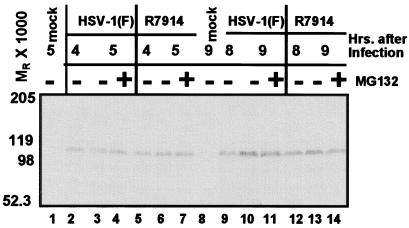
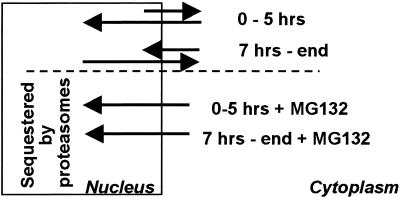
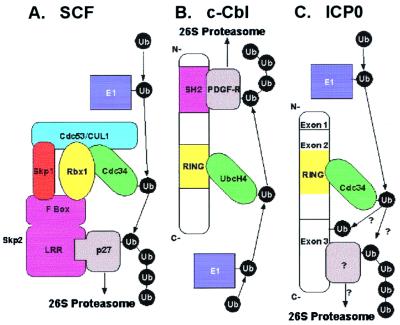
The infected cell protein 0 (ICP0) of herpes simplex virus 1, a promiscuous transactivator shown to enhance the expression of genes introduced into cells by infection or transfection, interacts with numerous cellular proteins and has been linked to the disruption of ND10 and degradation of several proteins. ICP0 contains a RING finger domain characteristic of a class of E3 ubiquitin ligases. We report that: (i) in infected cells, ICP0 interacts dynamically with proteasomes and is bound to proteasomes in the presence of the proteasome inhibitor MG132. Also in infected cells, cdc34, a polyubiquitinated E2 ubiquitin-conjugating enzyme, exhibits increased ICP0-dependent dynamic interaction with proteasomes. (ii) In an in vitro substrate-independent ubiquitination system, the RING finger domain encoded by exon 2 of ICP0 binds cdc34, whereas the carboxyl-terminal domain of ICP0 functions as an E3 ligase independent of the RING finger domain. The results indicate that ICP0 can act as a unimolecular E3 ubiquitin ligase and that it promotes ubiquitin-protein ligation and binds the E2 cdc34. It differs from other unimolecular E3 ligases in that the domain containing the RING finger binds E2, whereas the ligase activity maps to a different domain of the protein. The results also suggest that ICP0 shuttles between nucleus and cytoplasm as a function of its dynamic interactions with proteasomes.
Figures
Similar articles
-
Herpes simplex virus 1-infected cell protein 0 contains two E3 ubiquitin ligase sites specific for different E2 ubiquitin-conjugating enzymes.Proc Natl Acad Sci U S A. 2002 Jan 22;99(2):631-6. doi: 10.1073/pnas.022531599. Proc Natl Acad Sci U S A. 2002. PMID: 11805320 Free PMC article.
-
Herpes simplex virus 1 mutant in which the ICP0 HUL-1 E3 ubiquitin ligase site is disrupted stabilizes cdc34 but degrades D-type cyclins and exhibits diminished neurotoxicity.J Virol. 2003 Dec;77(24):13194-202. doi: 10.1128/jvi.77.24.13194-13202.2003. J Virol. 2003. PMID: 14645576 Free PMC article.
-
Characterization of the novel E3 ubiquitin ligase encoded in exon 3 of herpes simplex virus-1-infected cell protein 0.Proc Natl Acad Sci U S A. 2002 Jun 11;99(12):7889-94. doi: 10.1073/pnas.122246999. Proc Natl Acad Sci U S A. 2002. PMID: 12060736 Free PMC article.
-
Natural substrates of the proteasome and their recognition by the ubiquitin system.Curr Top Microbiol Immunol. 2002;268:137-74. doi: 10.1007/978-3-642-59414-4_6. Curr Top Microbiol Immunol. 2002. PMID: 12083004 Review.
-
Targeting E3 ubiquitin ligases for cancer therapy.Cancer Biol Ther. 2003 Nov-Dec;2(6):623-9. Cancer Biol Ther. 2003. PMID: 14688465 Review.
Cited by
-
Herpes simplex virus UL56 interacts with and regulates the Nedd4-family ubiquitin ligase Itch.Virol J. 2010 Aug 3;7:179. doi: 10.1186/1743-422X-7-179. Virol J. 2010. PMID: 20682038 Free PMC article.
-
Overexpression of promyelocytic leukemia protein precludes the dispersal of ND10 structures and has no effect on accumulation of infectious herpes simplex virus 1 or its proteins.J Virol. 2002 Sep;76(18):9355-67. doi: 10.1128/jvi.76.18.9355-9367.2002. J Virol. 2002. PMID: 12186918 Free PMC article.
-
Ability of the human cytomegalovirus IE1 protein to modulate sumoylation of PML correlates with its functional activities in transcriptional regulation and infectivity in cultured fibroblast cells.J Virol. 2004 Jun;78(12):6527-42. doi: 10.1128/JVI.78.12.6527-6542.2004. J Virol. 2004. PMID: 15163746 Free PMC article.
-
Corrigendum: Intrauterine Viral Infections: Impact of Inflammation on Fetal Neurodevelopment.Front Neurosci. 2021 Dec 10;15:817697. doi: 10.3389/fnins.2021.817697. eCollection 2021. Front Neurosci. 2021. PMID: 34955742 Free PMC article.
-
Role of ICP0 in the strategy of conquest of the host cell by herpes simplex virus 1.J Virol. 2004 Mar;78(5):2169-78. doi: 10.1128/jvi.78.5.2169-2178.2004. J Virol. 2004. PMID: 14963113 Free PMC article. Review. No abstract available.
References
-
- Roizman B, Knipe D M. In: Herpes Simplex Viruses and Their Replication, in Fields Virology. 4th Ed. Howley P, Knipe D M, editors. Philadelphia: Lippincott; 2001. , in press.
-
- Stow N D, Stow E C. J Gen Virol. 1986;67:2571–2585. - PubMed
-
- Maul G G, Guldner H H, Spivack J G. J Gen Virol. 1993;74:2679–2690. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
