

Somatic and germinal mobility of the RescueMu transposon in transgenic maize
- PMID: 11449053
- PMCID: PMC139541
- DOI: 10.1105/tpc.010002
Somatic and germinal mobility of the RescueMu transposon in transgenic maize
Abstract
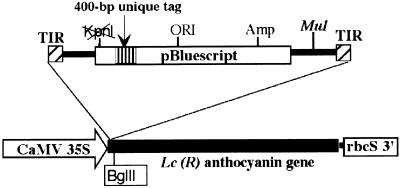
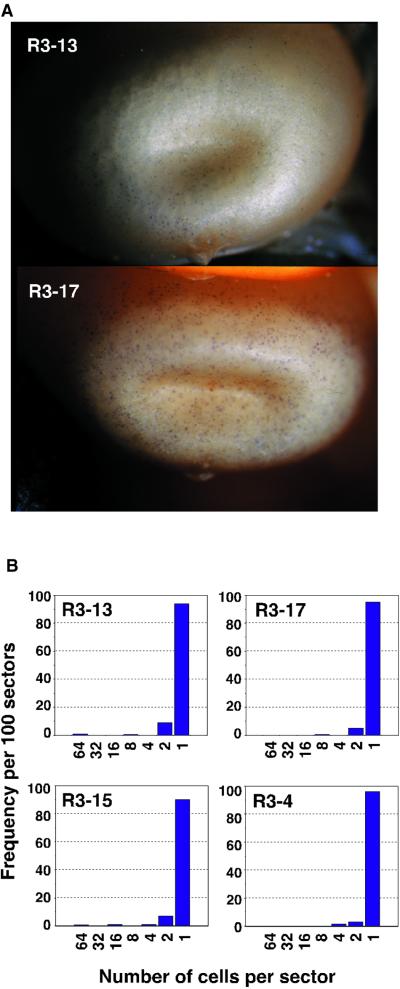
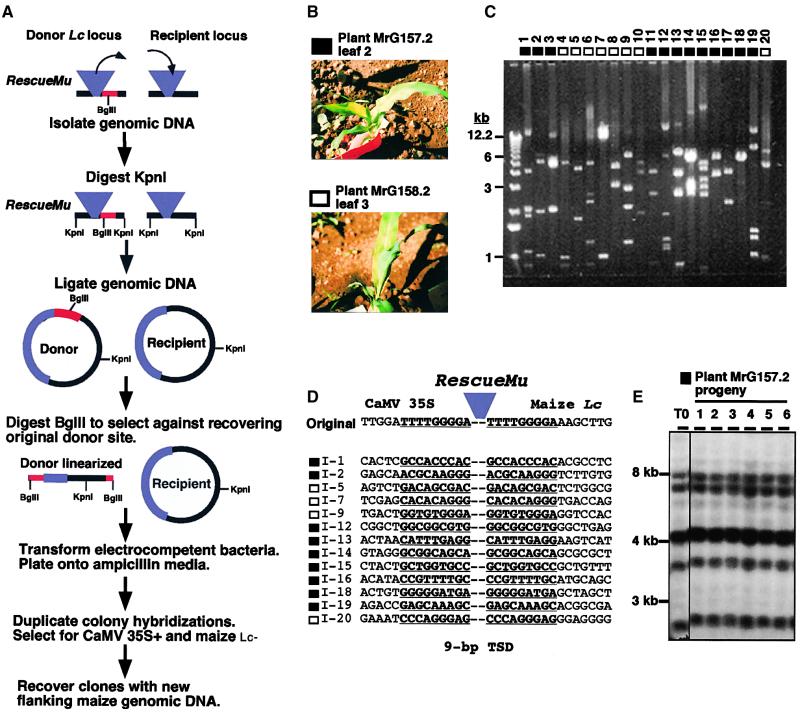
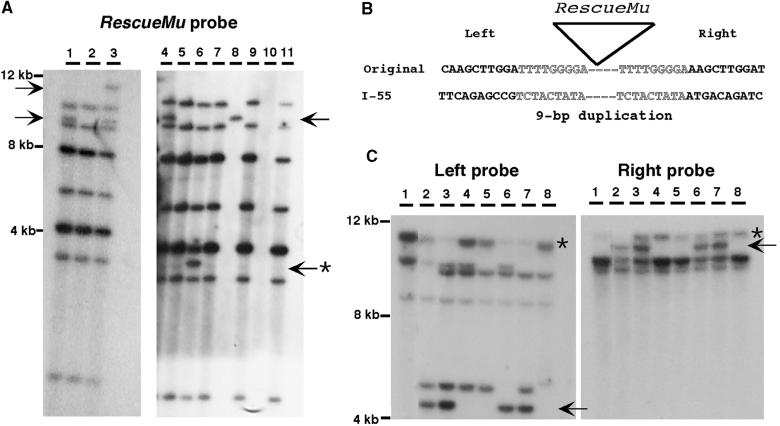
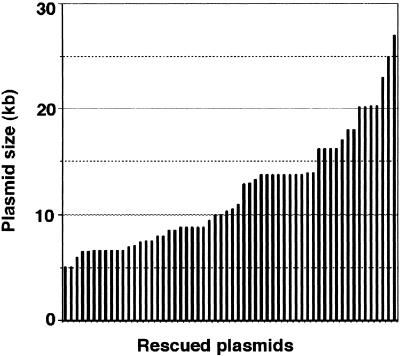
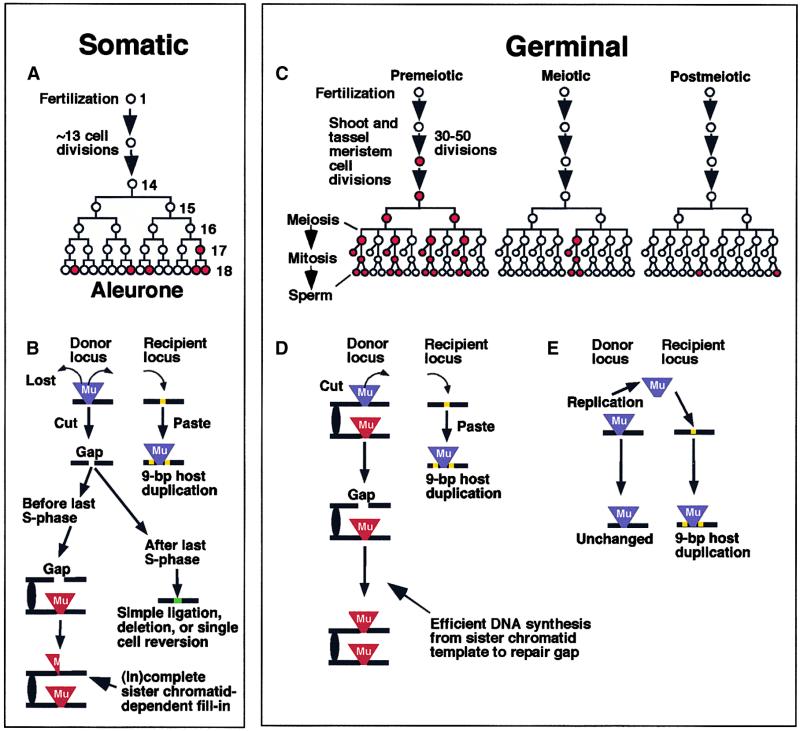
RescueMu, a Mu1 element containing a bacterial plasmid, is mobilized by MuDR in transgenic maize. Somatic excision from a cell-autonomous marker gene yields >90% single cell sectors; empty donor sites often have deletions and insertions, including up to 210 bp of RescueMu/Mu1 terminal DNA. Late somatic insertions are contemporaneous with excisions, suggesting that "cut-and-paste" transposition occurs in the soma. During reproduction, RescueMu transposes infrequently from the initial transgene array, but once transposed, RescueMu is suitable for high throughput gene mutation and cloning. As with MuDR/Mu elements, heritable RescueMu insertions are not associated with excisions. Both somatic and germinal RescueMu insertions occur preferentially into genes and gene-like sequences, but they exhibit weak target site preferences. New insights into Mu behaviors are discussed with reference to two models proposed to explain the alternative outcomes of somatic and germinal events: a switch from somatic cut-and-paste to germinal replicative transposition or to host-mediated gap repair from sister chromatids.
Figures

References
-
- Armstrong, C.L. (1994). Regeneration of plants from somatic cell cultures: Applications for in vitro genetic manipulation. In The Maize Handbook, M. Freeling and V. Walbot, eds (New York: Springer-Verlag), pp. 663–671.
-
- Armstrong, C.L., and Green, C.E. (1985). Establishment and maintenance of friable, embryogenic maize callus and the involvement of l-proline. Planta 164, 207–214. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
