

Topological and mutational analysis of Saccharomyces cerevisiae Ste14p, founding member of the isoprenylcysteine carboxyl methyltransferase family
- PMID: 11451995
- PMCID: PMC55642
- DOI: 10.1091/mbc.12.7.1957
Topological and mutational analysis of Saccharomyces cerevisiae Ste14p, founding member of the isoprenylcysteine carboxyl methyltransferase family
Abstract
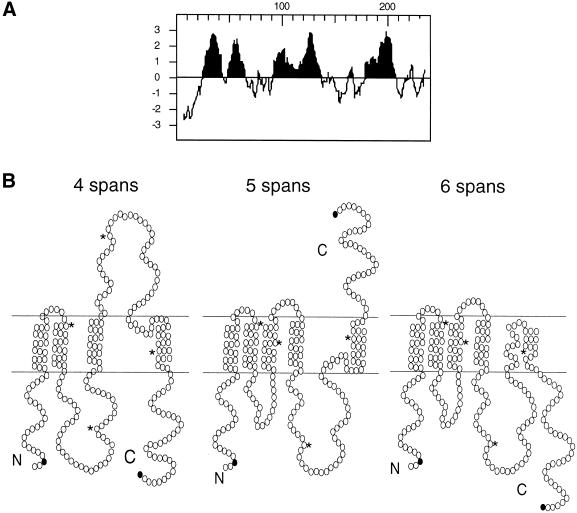
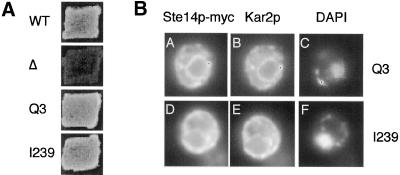
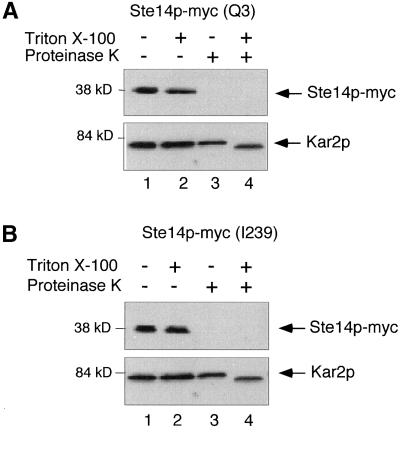
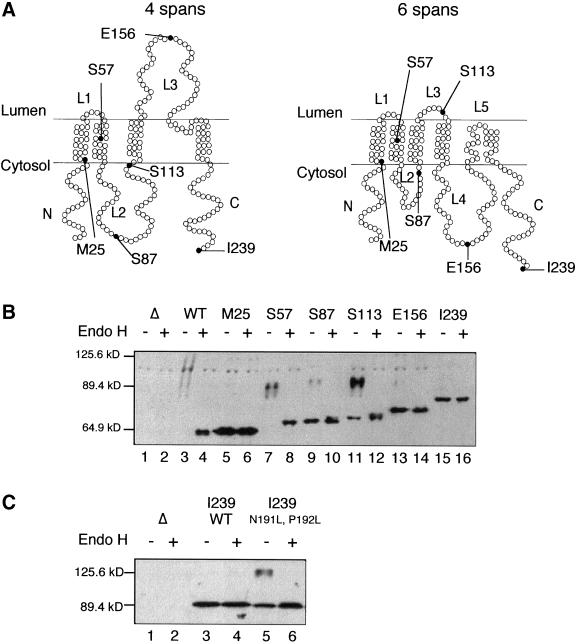
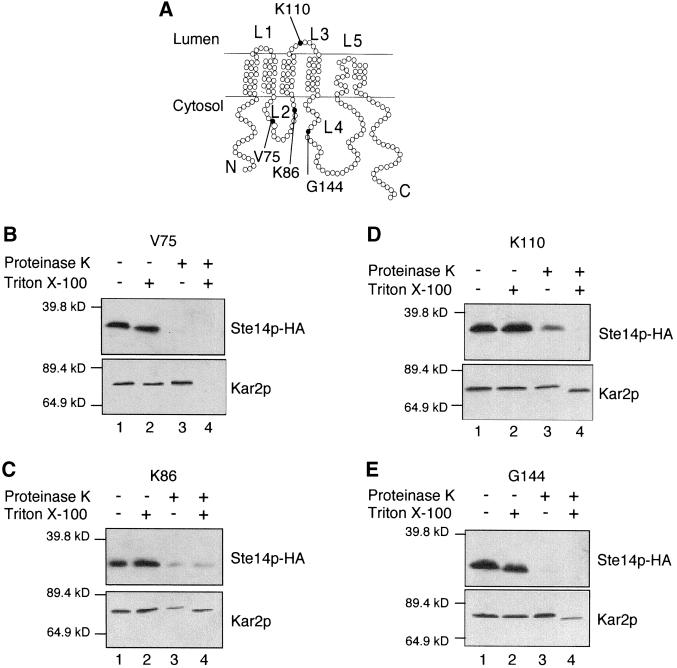
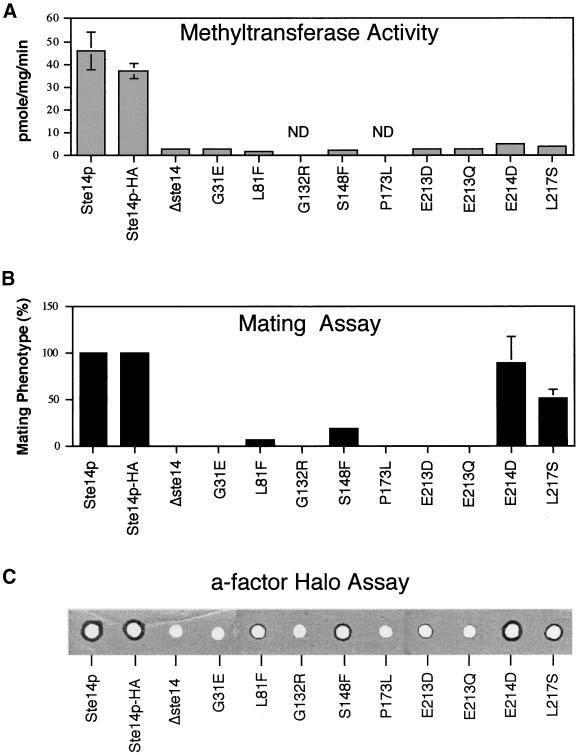
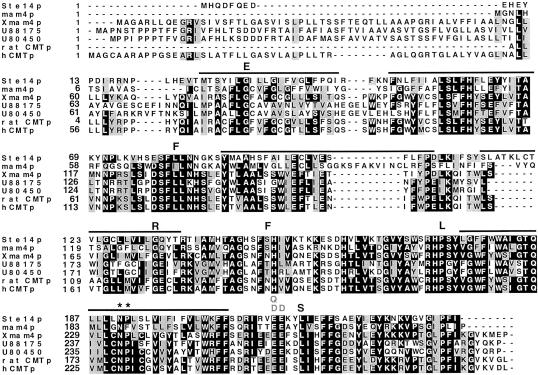
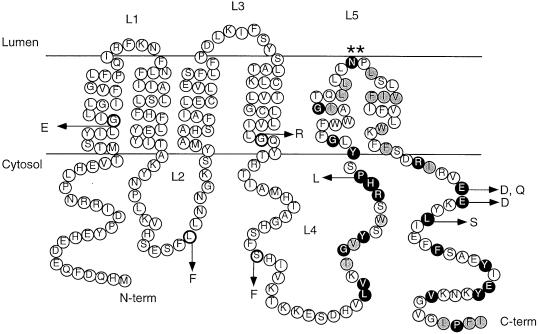
Eukaryotic proteins that terminate in a CaaX motif undergo three processing events: isoprenylation, C-terminal proteolytic cleavage, and carboxyl methylation. In Saccharomyces cerevisiae, the latter step is mediated by Ste14p, an integral endoplasmic reticulum membrane protein. Ste14p is the founding member of the isoprenylcysteine carboxyl methyltransferase (ICMT) family, whose members share significant sequence homology. Because the physiological substrates of Ste14p, such as Ras and the yeast a-factor precursor, are isoprenylated and reside on the cytosolic side of membranes, the Ste14p residues involved in enzymatic activity are predicted to be cytosolically disposed. In this study, we have investigated the topology of Ste14p by analyzing the protease protection of epitope-tagged versions of Ste14p and the glycosylation status of Ste14p-Suc2p fusions. Our data lead to a topology model in which Ste14p contains six membrane spans, two of which form a helical hairpin. According to this model most of the Ste14p hydrophilic regions are located in the cytosol. We have also generated ste14 mutants by random and site-directed mutagenesis to identify residues of Ste14p that are important for activity. Notably, four of the five loss-of-function mutations arising from random mutagenesis alter residues that are highly conserved among the ICMT family. Finally, we have identified a novel tripartite consensus motif in the C-terminal region of Ste14p. This region is similar among all ICMT family members, two phospholipid methyltransferases, several ergosterol biosynthetic enzymes, and a group of bacterial open reading frames of unknown function. Site-directed and random mutations demonstrate that residues in this region play a critical role in the function of Ste14p.
Figures

Similar articles
-
The Saccharomyces cerevisiae prenylcysteine carboxyl methyltransferase Ste14p is in the endoplasmic reticulum membrane.Mol Biol Cell. 1998 Aug;9(8):2231-47. doi: 10.1091/mbc.9.8.2231. Mol Biol Cell. 1998. PMID: 9693378 Free PMC article.
-
Functional oligomerization of the Saccharomyces cerevisiae isoprenylcysteine carboxyl methyltransferase, Ste14p.J Biol Chem. 2010 Apr 30;285(18):13380-7. doi: 10.1074/jbc.M109.061366. Epub 2010 Mar 3. J Biol Chem. 2010. PMID: 20202940 Free PMC article.
-
Purification, functional reconstitution, and characterization of the Saccharomyces cerevisiae isoprenylcysteine carboxylmethyltransferase Ste14p.J Biol Chem. 2005 Feb 25;280(8):7336-45. doi: 10.1074/jbc.M410292200. Epub 2004 Dec 20. J Biol Chem. 2005. PMID: 15611058
-
To finish things well: cysteine methylation ensures selective GTPase membrane localization and signalling.Curr Genet. 2018 Apr;64(2):341-344. doi: 10.1007/s00294-017-0756-x. Epub 2017 Sep 19. Curr Genet. 2018. PMID: 28929213 Review.
-
Carboxyl methylation of Ras-related proteins.Methods Enzymol. 1995;255:65-82. doi: 10.1016/s0076-6879(95)55010-0. Methods Enzymol. 1995. PMID: 8524139 Review. No abstract available.
Cited by
-
Evaluation of substrate and inhibitor binding to yeast and human isoprenylcysteine carboxyl methyltransferases (Icmts) using biotinylated benzophenone-containing photoaffinity probes.Biochem Biophys Res Commun. 2012 Jun 22;423(1):98-103. doi: 10.1016/j.bbrc.2012.05.089. Epub 2012 May 23. Biochem Biophys Res Commun. 2012. PMID: 22634004 Free PMC article.
-
A shunt pathway limits the CaaX processing of Hsp40 Ydj1p and regulates Ydj1p-dependent phenotypes.Elife. 2016 Aug 15;5:e15899. doi: 10.7554/eLife.15899. Elife. 2016. PMID: 27525482 Free PMC article.
-
Natural history of S-adenosylmethionine-binding proteins.BMC Struct Biol. 2005 Oct 14;5:19. doi: 10.1186/1472-6807-5-19. BMC Struct Biol. 2005. PMID: 16225687 Free PMC article.
-
Asparagine peptide lyases: a seventh catalytic type of proteolytic enzymes.J Biol Chem. 2011 Nov 4;286(44):38321-38328. doi: 10.1074/jbc.M111.260026. Epub 2011 Aug 8. J Biol Chem. 2011. PMID: 21832066 Free PMC article.
-
Uncovering the human methyltransferasome.Mol Cell Proteomics. 2011 Jan;10(1):M110.000976. doi: 10.1074/mcp.M110.000976. Epub 2010 Oct 7. Mol Cell Proteomics. 2011. PMID: 20930037 Free PMC article.
References
-
- Abeliovich H, Grote E, Novick P, Ferro-Novick S. Tlg2p, a yeast syntaxin homolog that resides on the Golgi and endocytic structures. J Biol Chem. 1998;273:11719–11727. - PubMed
-
- Ashby MN, Errada PR, Boyartchuk VL, Rine J. Isolation and DNA sequence of the STE14gene encoding farnesyl cysteine: carboxyl methyltransferase. Yeast. 1993;9:907–913. - PubMed
-
- Bergo MO, Leung GK, Ambroziak P, Otto JC, Casey PJ, Young SG. Targeted inactivation of the isoprenylcysteine carboxyl methyltransferase gene causes mislocalization of K-Ras in mammalian cells. J Biol Chem. 2000;275:17605–17610. - PubMed
-
- Berkower C. Analysis of STE6, a Saccharomyces cerevisiae ATP Binding Cassette (ABC) Protein. Ph.D. Thesis. Baltimore, MD: Johns Hopkins University; 1995.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
