

Telomere dysfunction triggers developmentally regulated germ cell apoptosis
- PMID: 11452000
- PMCID: PMC55650
- DOI: 10.1091/mbc.12.7.2023
Telomere dysfunction triggers developmentally regulated germ cell apoptosis
Abstract
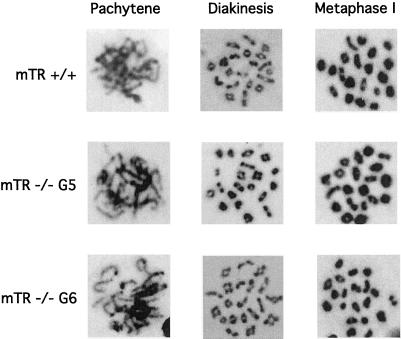
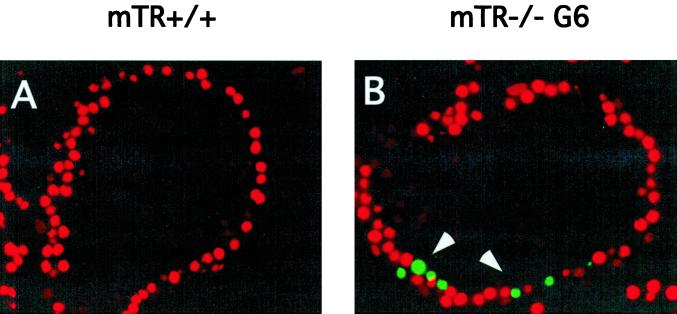
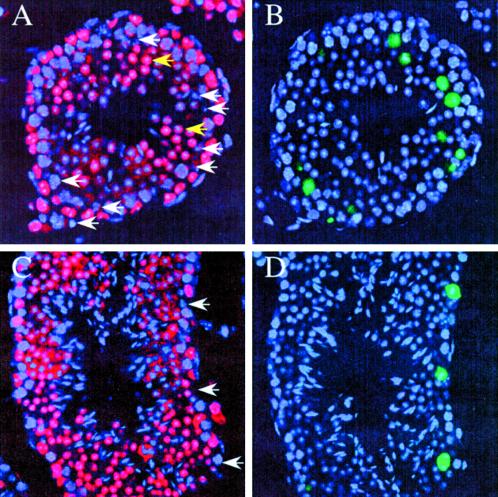
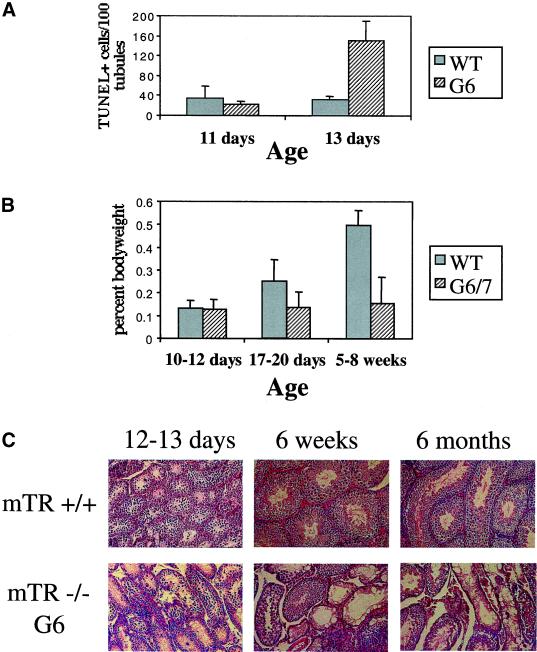
Telomere dysfunction results in fertility defects in a number of organisms. Although data from fission yeast and Caenorhabditis elegans suggests that telomere dysfunction manifests itself primarily as defects in proper meiotic chromosome segregation, it is unclear how mammalian telomere dysfunction results in germ cell death. To investigate the specific effects of telomere dysfunction on mammalian germ cell development, we examined the meiotic progression and germ cell apoptosis in late generation telomerase null mice. Our results indicate that chromosome asynapsis and missegregation are not the cause of infertility in mice with shortened telomeres. Rather, telomere dysfunction is recognized at the onset of meiosis, and cells with telomeric defects are removed from the germ cell precursor pool. This germ cell telomere surveillance may be an important mechanism to protect against the transmission of dysfunctional telomeres and chromosomal abnormalities.
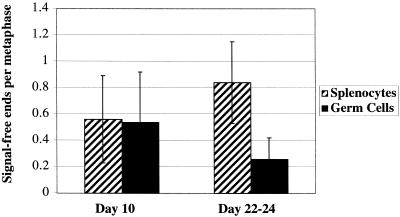
Figures
Similar articles
-
Telomere homeostasis in mammalian germ cells: a review.Chromosoma. 2016 Jun;125(2):337-51. doi: 10.1007/s00412-015-0555-4. Epub 2015 Nov 2. Chromosoma. 2016. PMID: 26525972 Review.
-
Irregular telomeres impair meiotic synapsis and recombination in mice.Proc Natl Acad Sci U S A. 2004 Apr 27;101(17):6496-501. doi: 10.1073/pnas.0400755101. Epub 2004 Apr 14. Proc Natl Acad Sci U S A. 2004. PMID: 15084742 Free PMC article.
-
Fission yeast Taz1 protein is required for meiotic telomere clustering and recombination.Nature. 1998 Apr 23;392(6678):828-31. doi: 10.1038/33947. Nature. 1998. PMID: 9572143
-
Telomeres and reproductive aging.Reprod Fertil Dev. 2009;21(1):10-4. doi: 10.1071/rd08229. Reprod Fertil Dev. 2009. PMID: 19152740 Review.
-
Mammalian Ku86 mediates chromosomal fusions and apoptosis caused by critically short telomeres.EMBO J. 2002 May 1;21(9):2207-19. doi: 10.1093/emboj/21.9.2207. EMBO J. 2002. PMID: 11980718 Free PMC article.
Cited by
-
Telomerase deficiency does not alter bleomycin-induced fibrosis in mice.Exp Lung Res. 2012 Apr;38(3):124-34. doi: 10.3109/01902148.2012.658148. Exp Lung Res. 2012. PMID: 22394286 Free PMC article.
-
Telomere homeostasis in mammalian germ cells: a review.Chromosoma. 2016 Jun;125(2):337-51. doi: 10.1007/s00412-015-0555-4. Epub 2015 Nov 2. Chromosoma. 2016. PMID: 26525972 Review.
-
High telomerase is a hallmark of undifferentiated spermatogonia and is required for maintenance of male germline stem cells.Genes Dev. 2015 Dec 1;29(23):2420-34. doi: 10.1101/gad.271783.115. Epub 2015 Nov 19. Genes Dev. 2015. PMID: 26584619 Free PMC article.
-
Endoreplication: polyploidy with purpose.Genes Dev. 2009 Nov 1;23(21):2461-77. doi: 10.1101/gad.1829209. Genes Dev. 2009. PMID: 19884253 Free PMC article. Review.
-
Irregular telomeres impair meiotic synapsis and recombination in mice.Proc Natl Acad Sci U S A. 2004 Apr 27;101(17):6496-501. doi: 10.1073/pnas.0400755101. Epub 2004 Apr 14. Proc Natl Acad Sci U S A. 2004. PMID: 15084742 Free PMC article.
References
-
- Ahmed S, Hodgkin J. MRT-2 checkpoint protein is required for germline immortality and telomere replication in C. elegans. Nature. 2000;403:159–164. - PubMed
-
- Allan DJ, Harmon BV, Kerr JFR. Cell death in spermatogenesis. In: Potten CS, editor. Perspectives on Mammalian Cell Death. London: Oxford University Press; 1987. pp. 229–258.
-
- Bellve AR. The molecular biology of mammalian spermatogenesis. In: Finn C A, editor. Oxford Reviews of Reproductive Biology. Oxford: Oxford University Press; 1979. pp. 159–261.
-
- Blasco MA, Lee HW, Hande MP, Samper E, Lansdorp PM, DePinho RA, Greider CW. Telomere shortening and tumor formation by mouse cells lacking telomerase RNA. Cell. 1997;91:25–34. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
