

Interleukin-12 primes CD4+ T cells for interferon-gamma production and protective immunity during Mycobacterium avium infection
- PMID: 11454066
- PMCID: PMC1783245
- DOI: 10.1046/j.1365-2567.2001.01237.x
Interleukin-12 primes CD4+ T cells for interferon-gamma production and protective immunity during Mycobacterium avium infection
Abstract
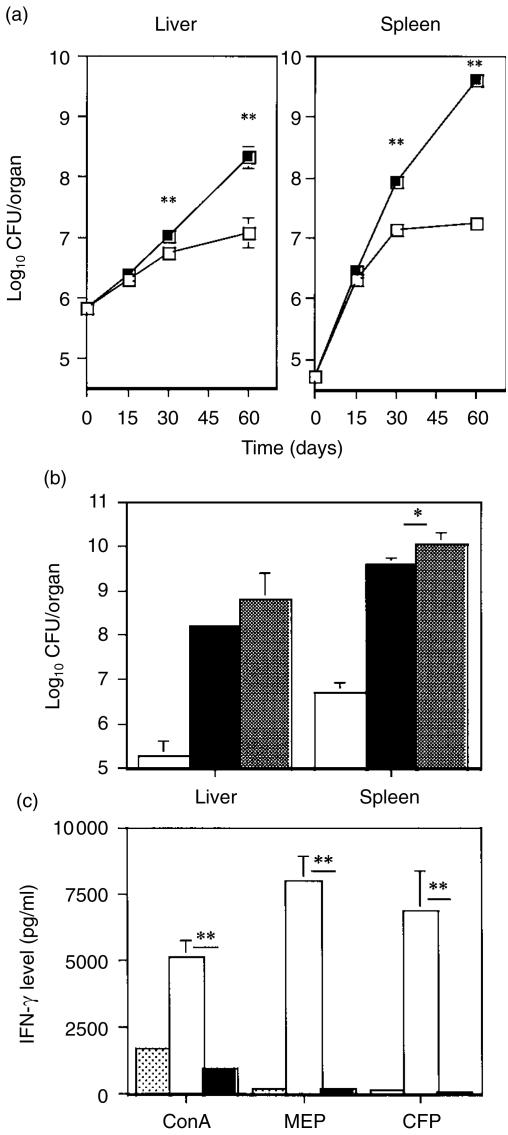
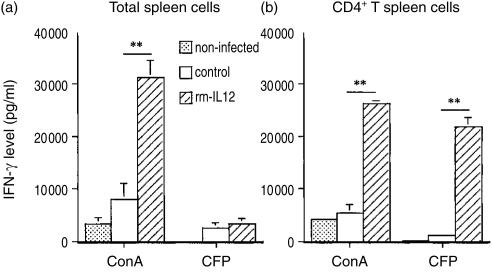
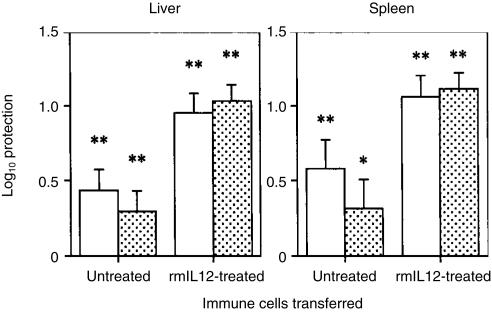
Interleukin-12 (IL-12) is a crucial cytokine for the generation of a protective immune response against Mycobacterium avium infection. In contrast to infected control mice, IL-12-deficient mice were unable to control bacterial proliferation and their spleen T cells were almost unresponsive in vitro to specific antigens of M. avium. Susceptibility of mice deficient in IL-12 was similar to that of interferon-gamma (IFN-gamma)-deficient mice. These data indicate a crucial role of IL-12 in the development of a T-cell population able to produce IFN-gamma and to mediate protection against M. avium infection. Treatment of M. avium-infected mice with IL-12 induced CD4+ T cells with enhanced capacity to produce IFN-gamma as well as to confer increased protection against M. avium.
Figures
Similar articles
-
Limited role of the Toll-like receptor-2 in resistance to Mycobacterium avium.Immunology. 2004 Feb;111(2):179-85. doi: 10.1111/j.0019-2805.2003.01807.x. Immunology. 2004. PMID: 15027903 Free PMC article.
-
The combination of plasmid interleukin-12 with a single DNA vaccine is more effective than Mycobacterium bovis (bacille Calmette-Guèrin) in protecting against systemic Mycobacterim avium infection.Immunology. 2003 Jun;109(2):308-14. doi: 10.1046/j.1365-2567.2003.01660.x. Immunology. 2003. PMID: 12757627 Free PMC article.
-
Protection against virulent Mycobacterium avium infection following DNA vaccination with the 35-kilodalton antigen is accompanied by induction of gamma interferon-secreting CD4(+) T cells.Infect Immun. 2000 Jun;68(6):3090-6. doi: 10.1128/IAI.68.6.3090-3096.2000. Infect Immun. 2000. PMID: 10816448 Free PMC article.
-
Anti-CD3 single-chain Fv/interleukin-18 fusion DNA induces anti-mycobacterial resistance via efficient interferon-gamma production in BALB/c mice infected with Mycobacterium avium.Vaccine. 2006 Apr 12;24(16):3365-73. doi: 10.1016/j.vaccine.2005.12.065. Epub 2006 Jan 31. Vaccine. 2006. PMID: 16481075
-
Protective role of interferon gamma, tumor necrosis factor alpha and interleukin-6 in Mycobacterium tuberculosis and M. avium infections.Immunobiology. 1994 Oct;191(4-5):520-5. doi: 10.1016/S0171-2985(11)80458-4. Immunobiology. 1994. PMID: 7713566 Review.
Cited by
-
BCG Vaccination Induces M. avium and M. abscessus Cross-Protective Immunity.Front Immunol. 2019 Feb 19;10:234. doi: 10.3389/fimmu.2019.00234. eCollection 2019. Front Immunol. 2019. PMID: 30837992 Free PMC article.
-
Mycobacterium avium Complex Disease.Microbiol Spectr. 2017 Apr;5(2):10.1128/microbiolspec.tnmi7-0045-2017. doi: 10.1128/microbiolspec.TNMI7-0045-2017. Microbiol Spectr. 2017. PMID: 28429679 Free PMC article. Review.
-
Heme catabolism by heme oxygenase-1 confers host resistance to Mycobacterium infection.Infect Immun. 2013 Jul;81(7):2536-45. doi: 10.1128/IAI.00251-13. Epub 2013 Apr 29. Infect Immun. 2013. PMID: 23630967 Free PMC article.
-
Inherited interleukin-12 deficiency: IL12B genotype and clinical phenotype of 13 patients from six kindreds.Am J Hum Genet. 2002 Feb;70(2):336-48. doi: 10.1086/338625. Epub 2001 Dec 17. Am J Hum Genet. 2002. PMID: 11753820 Free PMC article.
-
Nontuberculous mycobacterial osteomyelitis.Infect Dis (Lond). 2015;47(10):673-85. doi: 10.3109/23744235.2015.1040445. Epub 2015 Apr 27. Infect Dis (Lond). 2015. PMID: 25915177 Free PMC article. Review.
References
-
- Wolinsky E. Mycobacterial diseases other than tuberculosis. Clin Infect Dis. 1992;15:1–10. - PubMed
-
- Wolinsky E. Mycobacterial lymphadenitis in children: a prospective study of 105 nontuberculous cases with long-term follow-up. Clin Infect Dis. 1995;20:954–63. - PubMed
-
- Prince DS, Peterson DD, Steiner RM, Gottlieb JE, Scott R, Israel HL, Figueroa WG, Fish JE. Infection with Mycobacterium avium complex in patients without predisposing conditions. N Engl J Med. 1989;321:863–8. - PubMed
-
- Horsburgh CR. Mycobacterium avium complex infection in the acquired immunodeficiency syndrome. N Engl J Med. 1991;324:1332–8. - PubMed
-
- Jacobson MA, Hopewell PC, Yajko DM, et al. Natural history of disseminated Mycobacterium avium complex infection in AIDS. J Infect Dis. 1991;164:994–8. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
