

Serotonin facilitates AMPA-type responses in isolated siphon motor neurons of Aplysia in culture
- PMID: 11454967
- PMCID: PMC2278700
- DOI: 10.1111/j.1469-7793.2001.00501.x
Serotonin facilitates AMPA-type responses in isolated siphon motor neurons of Aplysia in culture
Abstract
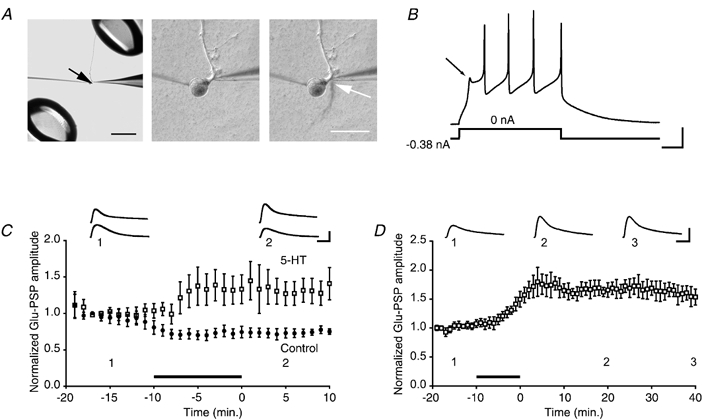
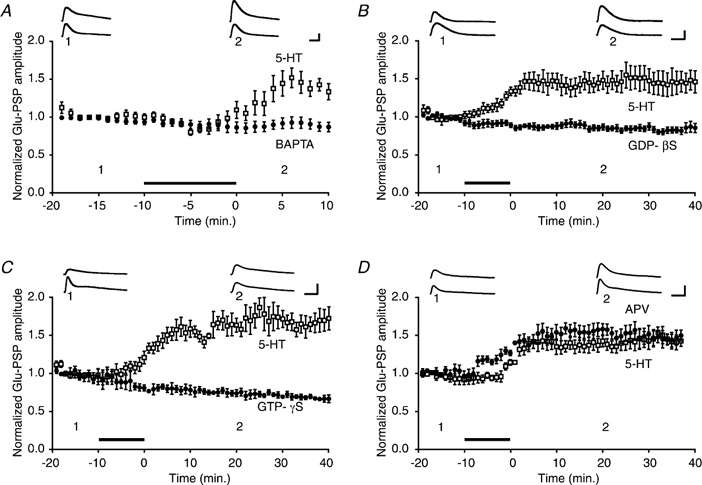
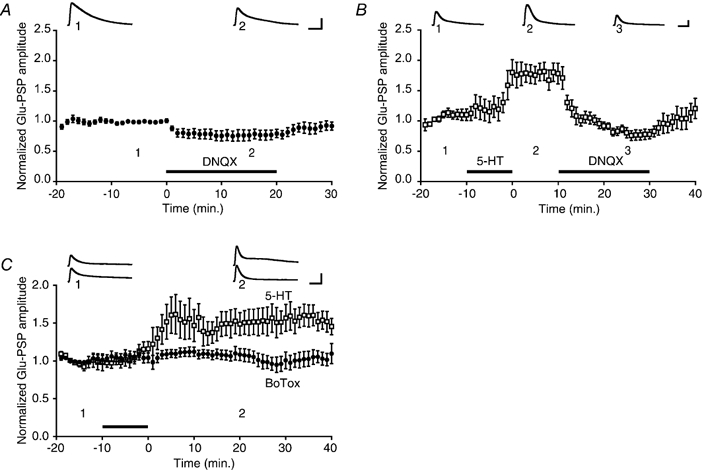
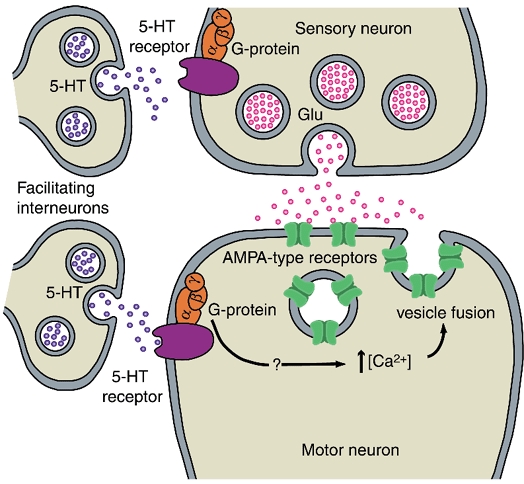
1. Serotonin (5-HT) facilitates the connections between sensory and motor neurons in Aplysia during behavioural sensitization. The effect of 5-HT on sensorimotor synapses is believed to be primarily presynaptic. Here we tested whether 5-HT can have an exclusively postsynaptic facilitatory effect. 2. Siphon motor neurons were individually dissociated from the abdominal ganglion of Aplysia and placed into cell culture. Brief pulses of glutamate, the putative sensory neuron transmitter, were focally applied (0.1 Hz) to solitary motor neurons in culture, and the glutamate-evoked postsynaptic potentials (Glu-PSPs) were recorded. 3. When 5-HT was perfused over the motor neuron for 10 min, the amplitude of the Glu-PSPs was significantly increased. The 5-HT-induced enhancement of the Glu-PSPs persisted for at least 40 min after washout. 4. Prior injection into the motor neuron of the calcium chelator BAPTA, GDP-beta-S or GTP-gamma-S blocked the 5-HT-induced facilitation of the Glu-PSPs. However, the facilitation was not blocked when APV, an NMDA receptor antagonist, was applied together with the 5-HT. 5. The enhancement of the Glu-PSPs by 5-HT was reversed by the AMPA receptor antagonist DNQX, indicating that 5-HT increased the functional expression of AMPA-type receptors in the motor neuron. 6. The presence of botulinum toxin in the motor neuron blocked the 5-HT-induced enhancement of the Glu-PSPs. As botulinum toxin prevents exocytosis we hypothesize that during sensitization 5-HT causes the insertion of additional AMPA-type receptors into the postsynaptic membrane of sensorimotor synapses via exocytosis. This postsynaptic mechanism may contribute to facilitation of the synapses.
Figures
Similar articles
-
Synaptic facilitation and behavioral dishabituation in Aplysia: dependence on release of Ca2+ from postsynaptic intracellular stores, postsynaptic exocytosis, and modulation of postsynaptic AMPA receptor efficacy.J Neurosci. 2005 Jun 8;25(23):5623-37. doi: 10.1523/JNEUROSCI.5305-04.2005. J Neurosci. 2005. PMID: 15944390 Free PMC article.
-
The potential role of postsynaptic phospholipase C activity in synaptic facilitation and behavioral sensitization in Aplysia.J Neurophysiol. 2008 Jul;100(1):108-16. doi: 10.1152/jn.90389.2008. Epub 2008 May 14. J Neurophysiol. 2008. PMID: 18480365 Free PMC article.
-
Involvement of presynaptic and postsynaptic mechanisms in a cellular analog of classical conditioning at Aplysia sensory-motor neuron synapses in isolated cell culture.J Neurosci. 1998 Jan 1;18(1):458-66. doi: 10.1523/JNEUROSCI.18-01-00458.1998. J Neurosci. 1998. PMID: 9412522 Free PMC article.
-
Postsynaptic regulation of the development and long-term plasticity of Aplysia sensorimotor synapses in cell culture.J Neurobiol. 1994 Jun;25(6):666-93. doi: 10.1002/neu.480250608. J Neurobiol. 1994. PMID: 8071666 Review.
-
Habituation in Aplysia: the Cheshire cat of neurobiology.Neurobiol Learn Mem. 2009 Sep;92(2):147-54. doi: 10.1016/j.nlm.2009.03.005. Epub 2009 Mar 28. Neurobiol Learn Mem. 2009. PMID: 19332142 Review.
Cited by
-
Homolog of protein kinase Mζ maintains context aversive memory and underlying long-term facilitation in terrestrial snail Helix.Front Cell Neurosci. 2015 Jun 22;9:222. doi: 10.3389/fncel.2015.00222. eCollection 2015. Front Cell Neurosci. 2015. PMID: 26157359 Free PMC article.
-
Role of nitric oxide in classical conditioning of siphon withdrawal in Aplysia.J Neurosci. 2007 Oct 10;27(41):10993-1002. doi: 10.1523/JNEUROSCI.2357-07.2007. J Neurosci. 2007. PMID: 17928440 Free PMC article.
-
Facilitation of monosynaptic and complex PSPs in type I interneurons of conditioned Hermissenda.J Neurosci. 2002 Sep 1;22(17):7818-24. doi: 10.1523/JNEUROSCI.22-17-07818.2002. J Neurosci. 2002. PMID: 12196605 Free PMC article.
-
Co-induction of LTP and LTD and its regulation by protein kinases and phosphatases.J Neurophysiol. 2010 May;103(5):2737-46. doi: 10.1152/jn.01112.2009. Epub 2010 Mar 24. J Neurophysiol. 2010. PMID: 20457859 Free PMC article.
-
The shift of the reversal potential of glutamate-activated chloride current in molluscan neurons caused by 8-Br-cAMP.Dokl Biol Sci. 2004 Nov-Dec;399:447-50. doi: 10.1007/s10630-005-0008-y. Dokl Biol Sci. 2004. PMID: 15717604 No abstract available.
References
-
- Berridge MJ. Inositol trisphosphate and calcium signaling. Nature. 1993;361:315–325. - PubMed
-
- Chitwood RA, Li Q, Glanzman DL. Serotonin enhances the glutamate response in isolated Aplysia motor neurons in culture. Society for Neuroscience Abstracts. 2000;26:1526.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials