

Class I beta-1,3-glucanase and chitinase are expressed in the micropylar endosperm of tomato seeds prior to radicle emergence
- PMID: 11457981
- PMCID: PMC116487
- DOI: 10.1104/pp.126.3.1299
Class I beta-1,3-glucanase and chitinase are expressed in the micropylar endosperm of tomato seeds prior to radicle emergence
Abstract
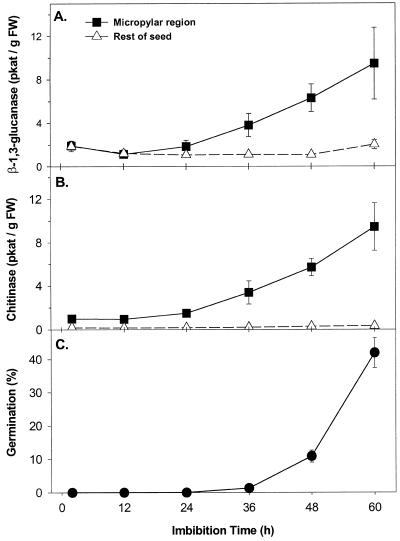
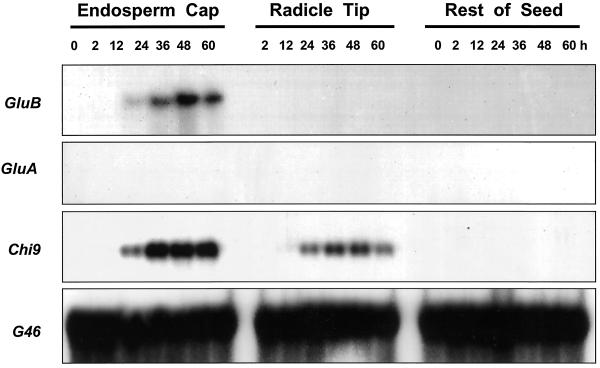
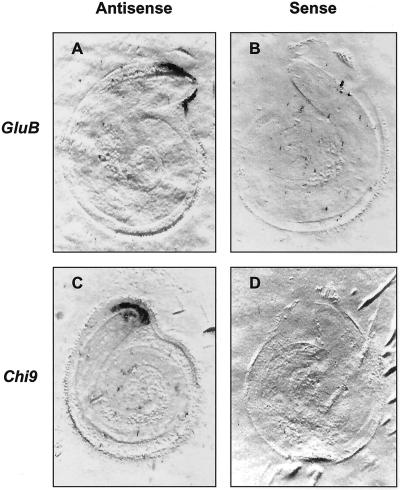
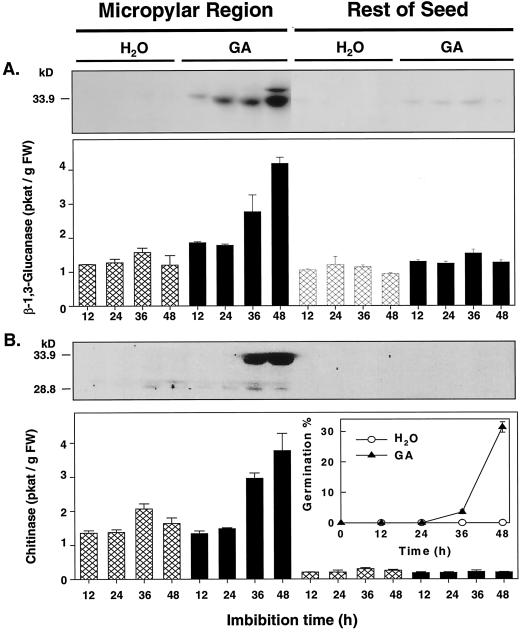
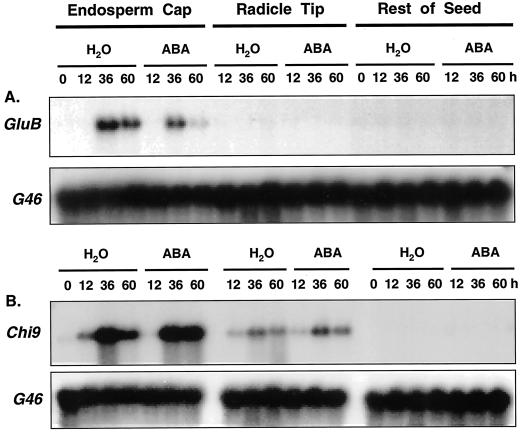
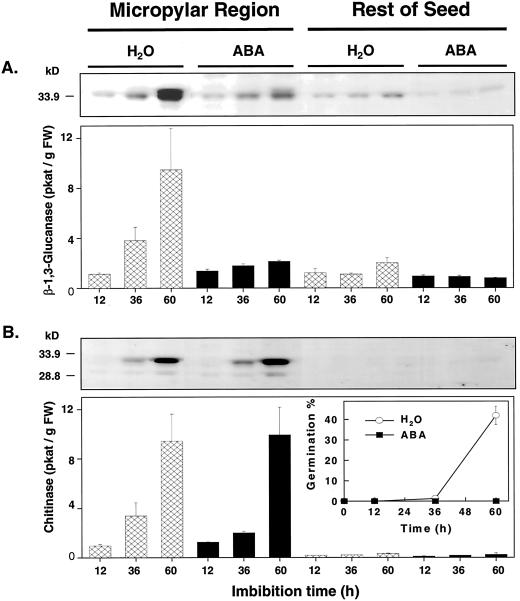
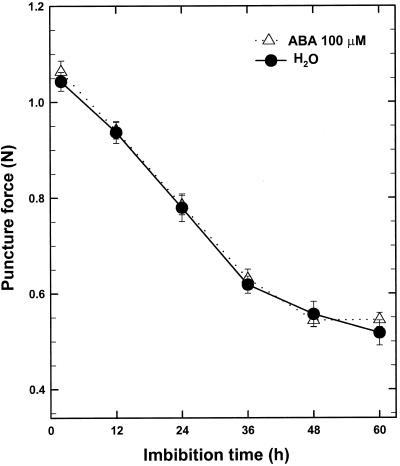
beta-1,3-Glucanase (EC 3.2.1.39) and chitinase (EC 3.2.1.14) mRNAs, proteins, and enzyme activities were expressed specifically in the micropylar tissues of imbibed tomato (Lycopersicon esculentum Mill.) seeds prior to radicle emergence. RNA hybridization and immunoblotting demonstrated that both enzymes were class I basic isoforms. beta-1,3-Glucanase was expressed exclusively in the endosperm cap tissue, whereas chitinase localized to both endosperm cap and radicle tip tissues. beta-1,3-Glucanase and chitinase appeared in the micropylar tissues of gibberellin-deficient gib-1 tomato seeds only when supplied with gibberellin. Accumulation of beta-1,3-glucanase mRNA, protein and enzyme activity was reduced by 100 microM abscisic acid, which delayed or prevented radicle emergence but not endosperm cap weakening. In contrast, expression of chitinase mRNA, protein, and enzyme activity was not affected by abscisic acid. Neither of these enzymes significantly hydrolyzed isolated tomato endosperm cap cell walls. Although both beta-1,3-glucanase and chitinase were expressed in tomato endosperm cap tissue prior to radicle emergence, we found no evidence that they were directly involved in cell wall modification or tissue weakening. Possible functions of these hydrolases during tomato seed germination are discussed.
Figures

Similar articles
-
A germination-specific endo-beta-mannanase gene is expressed in the micropylar endosperm cap of tomato seeds.Plant Physiol. 2000 Aug;123(4):1235-46. doi: 10.1104/pp.123.4.1235. Plant Physiol. 2000. PMID: 10938343 Free PMC article.
-
Class I chitinase and beta-1,3-glucanase are differentially regulated by wounding, methyl jasmonate, ethylene, and gibberellin in tomato seeds and leaves.Plant Physiol. 2003 Sep;133(1):263-73. doi: 10.1104/pp.103.024687. Plant Physiol. 2003. PMID: 12970492 Free PMC article.
-
Expression of an expansin is associated with endosperm weakening during tomato seed germination.Plant Physiol. 2000 Nov;124(3):1265-74. doi: 10.1104/pp.124.3.1265. Plant Physiol. 2000. PMID: 11080302 Free PMC article.
-
Induction of α-amylase and endosperm-imposed seed dormancy: two pioneering papers in gibberellin research.Planta. 2025 Apr 25;261(6):118. doi: 10.1007/s00425-025-04699-w. Planta. 2025. PMID: 40278915 Free PMC article. Review.
-
The biomechanics of seed germination.J Exp Bot. 2017 Feb 1;68(4):765-783. doi: 10.1093/jxb/erw428. J Exp Bot. 2017. PMID: 27927995 Review.
Cited by
-
A study of some biochemical and histopathological responses of wet-stored recalcitrant seeds of Avicennia marina infected by Fusarium moniliforme.Ann Bot. 2003 Sep;92(3):401-8. doi: 10.1093/aob/mcg154. Epub 2003 Jul 24. Ann Bot. 2003. PMID: 12881405 Free PMC article.
-
The perisperm-endosperm envelope in Cucumis: structure, proton diffusion and cell wall hydrolysing activity.Ann Bot. 2005 Oct;96(5):769-78. doi: 10.1093/aob/mci234. Epub 2005 Jul 25. Ann Bot. 2005. PMID: 16043437 Free PMC article.
-
Mechanism and control of Solanum lycocarpum seed germination.Ann Bot. 2007 Dec;100(6):1175-87. doi: 10.1093/aob/mcm211. Epub 2007 Sep 11. Ann Bot. 2007. PMID: 17855380 Free PMC article.
-
Expression of cell wall related genes in basal and ear internodes of silking brown-midrib-3, caffeic acid O-methyltransferase (COMT) down-regulated, and normal maize plants.BMC Plant Biol. 2008 Jun 26;8:71. doi: 10.1186/1471-2229-8-71. BMC Plant Biol. 2008. PMID: 18582385 Free PMC article.
-
Expression analysis of chitinase upon challenge inoculation to Alternaria wounding and defense inducers in Brassica juncea.Biotechnol Rep (Amst). 2017 Jan 5;13:72-79. doi: 10.1016/j.btre.2017.01.001. eCollection 2017 Mar. Biotechnol Rep (Amst). 2017. PMID: 28352565 Free PMC article.
References
-
- Albersheim P, Nevins DJ, English PD, Karr A. A method for analysis of sugars in plant cell wall polysaccharides by GLC. Carbohydr Res. 1967;5:340–345.
-
- Ausubel FM, Brent R, Kinston RE, Moore DD, Smith JA, Seidman JG, Struhl K. Current Protocols in Molecular Biology. New York: Wiley-Interscience; 1987.
-
- Ballance GM, Meredith WOS, Laberge DE. Distribution and development of endo-β-glucanase activities in barley tissues during germination. Can J Plant Sci. 1976;56:459–466.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
