

Messenger RNAs are recruited for nuclear export during transcription
- PMID: 11459827
- PMCID: PMC312744
- DOI: 10.1101/gad.892401
Messenger RNAs are recruited for nuclear export during transcription
Abstract
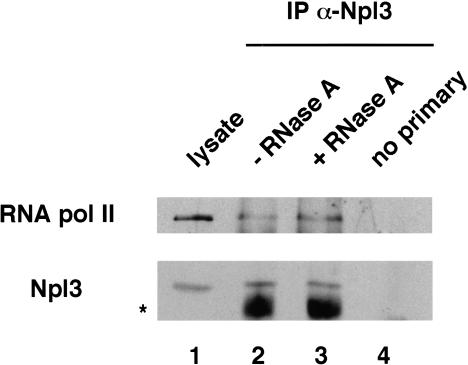
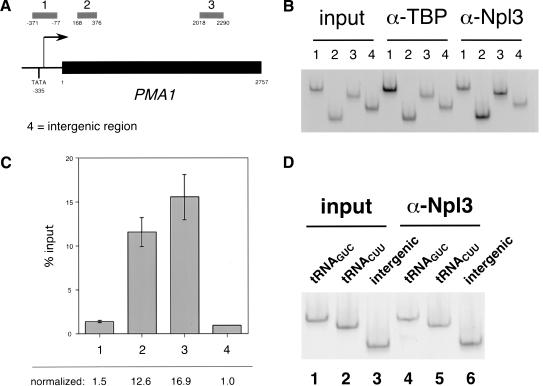
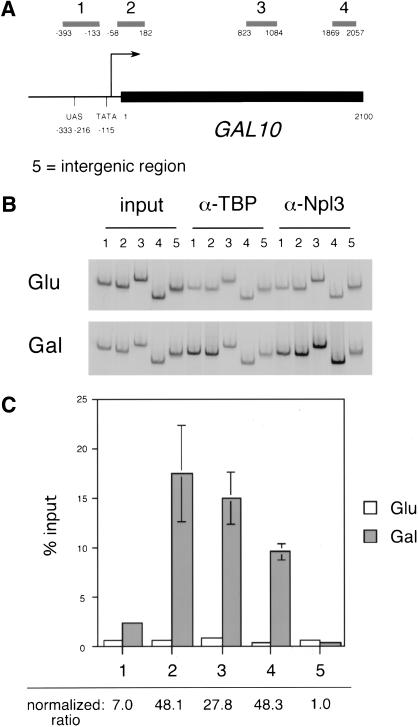
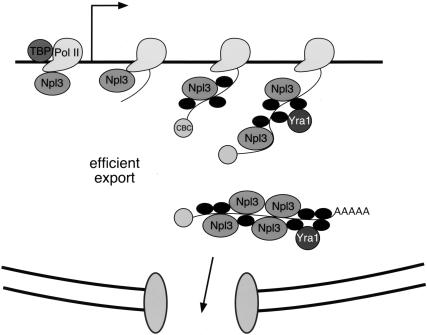
Following transcription and processing, eukaryotic mRNAs are exported from the nucleus to the cytoplasm for translation. Here we present evidence that mRNAs are targeted for nuclear export cotranscriptionally. Combined mutations in the Saccharomyces cerevisiae hnRNP Npl3 and TATA-binding protein (TBP) block mRNA export, implying that cotranscriptional recruitment of Npl3 is required for efficient export of mRNA. Furthermore, Npl3 can be found in a complex with RNA Pol II, indicating that Npl3 associates with the transcription machinery. Finally, Npl3 is recruited to genes in a transcription dependent manner as determined by chromatin immunoprecipitation. Another mRNA export factor, Yra1, also associates with chromatin cotranscriptionally but appears to be recruited at a later step. Taken together, our results suggest that export factors are recruited to the sites of transcription to promote efficient mRNA export.
Figures




References
-
- Alzhanova-Ericsson AT, Sun X, Visa N, Kiseleva E, Wurtz T, Daneholt B. A protein of the SR family of splicing factors binds extensively to exonic Balbiani ring pre-mRNA and accompanies the RNA from the gene to the nuclear pore. Genes & Dev. 1996;10:2881–2893. - PubMed
-
- Amberg DC, Goldstein AL, Cole CN. Isolation and characterization of RAT1: An essential gene of Saccharomyces cerevisiae required for the efficient nucleocytoplasmic trafficking of mRNA. Genes & Dev. 1992;6:1173–1189. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials