

Rescue of arrested replication forks by homologous recombination
- PMID: 11459951
- PMCID: PMC37419
- DOI: 10.1073/pnas.111008798
Rescue of arrested replication forks by homologous recombination
Abstract
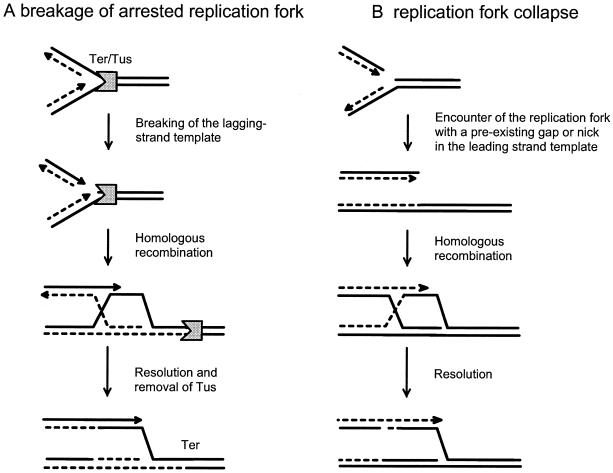
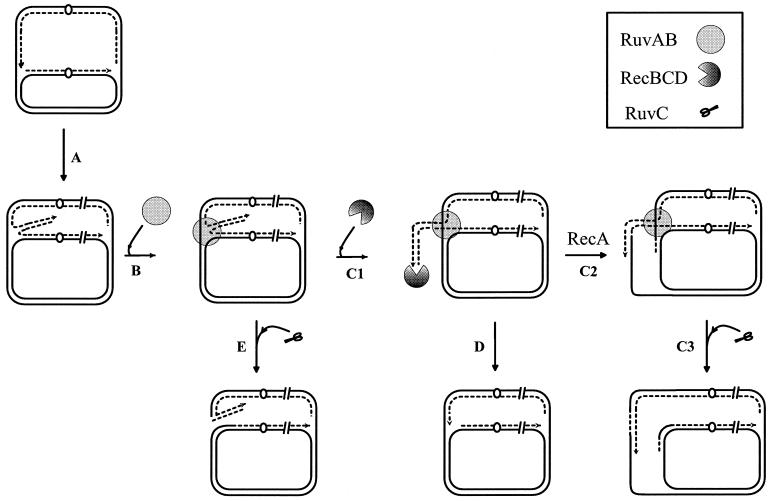
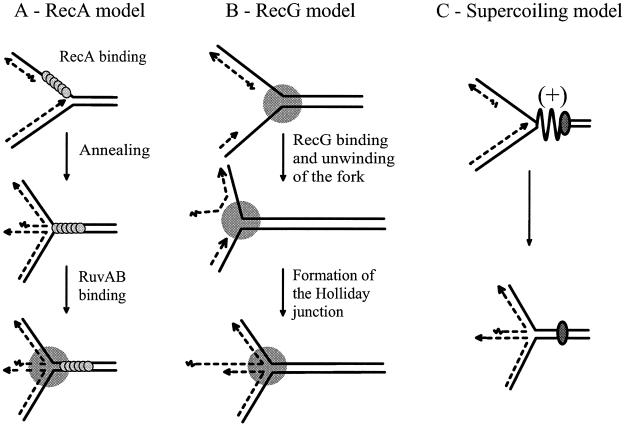
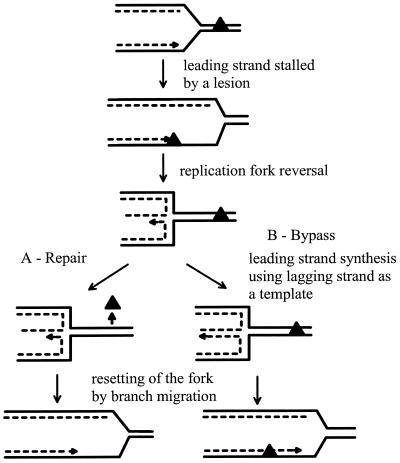
DNA synthesis is an accurate and very processive phenomenon; nevertheless, replication fork progression on chromosomes can be impeded by DNA lesions, DNA secondary structures, or DNA-bound proteins. Elements interfering with the progression of replication forks have been reported to induce rearrangements and/or render homologous recombination essential for viability, in all organisms from bacteria to human. Arrested replication forks may be the target of nucleases, thereby providing a substrate for double-strand break repair enzyme. For example in bacteria, direct fork breakage was proposed to occur at replication forks blocked by a bona fide replication terminator sequence, a specific site that arrests bacterial chromosome replication. Alternatively, an arrested replication fork may be transformed into a recombination substrate by reversal of the forked structures. In reversed forks, the last duplicated portions of the template strands reanneal, allowing the newly synthesized strands to pair. In bacteria, this reaction was proposed to occur in replication mutants, in which fork arrest is caused by a defect in a replication protein, and in UV irradiated cells. Recent studies suggest that it may also occur in eukaryote organisms. We will review here observations that link replication hindrance with DNA rearrangements and the possible underlying molecular processes.
Figures
References
-
- Seitz E M, Kowalczykowski S C. Mol Microbiol. 2000;37:555–560. - PubMed
-
- Egelman E. Trends Biochem Sci. 2000;25:180–184. - PubMed
-
- Michel B. In: Organization of the Prokaryotic Genome. Charlebois R L, editor. Washington, DC: Am. Soc. Microbiol.; 1999. pp. 129–150.
-
- Lewis L K, Resnick M A. Mutat Res. 2000;451:71–89. - PubMed
-
- Lieber M R. Genes Cells. 1999;4:77–85. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
