

Break-induced replication: a review and an example in budding yeast
- PMID: 11459961
- PMCID: PMC37429
- DOI: 10.1073/pnas.151008198
Break-induced replication: a review and an example in budding yeast
Abstract
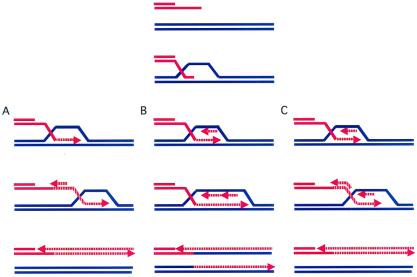
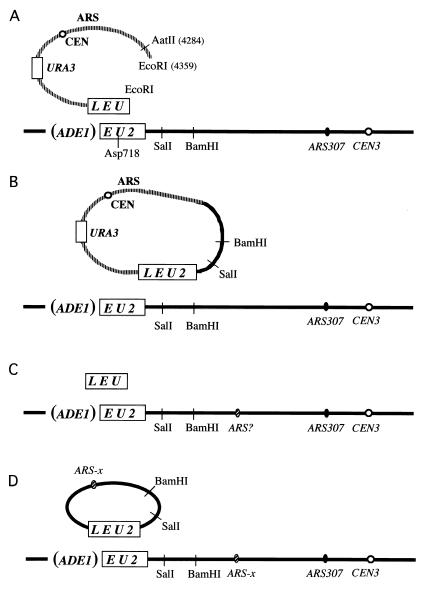
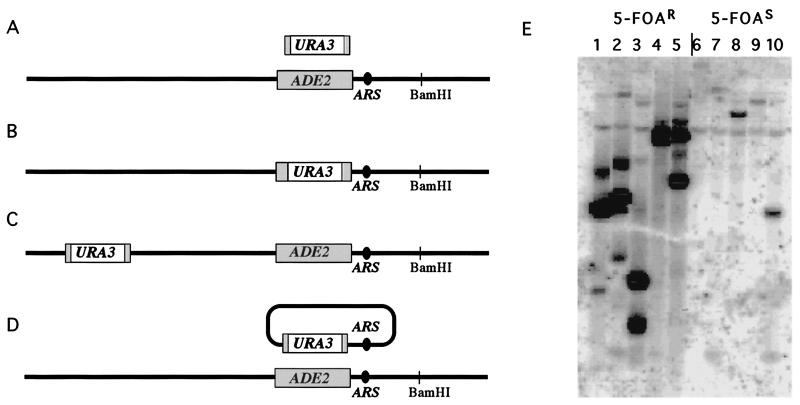
Break-induced replication (BIR) is a nonreciprocal recombination-dependent replication process that is an effective mechanism to repair a broken chromosome. We review key roles played by BIR in maintaining genome integrity, including restarting DNA replication at broken replication forks and maintaining telomeres in the absence of telomerase. Previous studies suggested that gene targeting does not occur by simple crossings-over between ends of the linearized transforming fragment and the target chromosome, but involves extensive new DNA synthesis resembling BIR. We examined gene targeting in Saccharomyces cerevisiae where only one end of the transformed DNA has homology to chromosomal sequences. Linearized, centromere-containing plasmid DNA with the 5' end of the LEU2 gene at one end was transformed into a strain in which the 5' end of LEU2 was replaced by ADE1, preventing simple homologous gene replacement to become Leu2(+). Ade1(+) Leu2(+) transformants were recovered in which the entire LEU2 gene and as much as 7 kb of additional sequences were found on the plasmid, joined by microhomologies characteristic of nonhomologous end-joining (NHEJ). In other experiments, cells were transformed with DNA fragments lacking an ARS and homologous to only 50 bp of ADE2 added to the ends of a URA3 gene. Autonomously replicating circles were recovered, containing URA3 and as much as 8 kb of ADE2-adjacent sequences, including a nearby ARS, copied from chromosomal DNA. Thus, the end of a linearized DNA fragment can initiate new DNA synthesis by BIR in which the newly synthesized DNA is displaced and subsequently forms circles by NHEJ.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
