

Rad54 protein stimulates the postsynaptic phase of Rad51 protein-mediated DNA strand exchange
- PMID: 11459988
- PMCID: PMC37456
- DOI: 10.1073/pnas.121009898
Rad54 protein stimulates the postsynaptic phase of Rad51 protein-mediated DNA strand exchange
Abstract
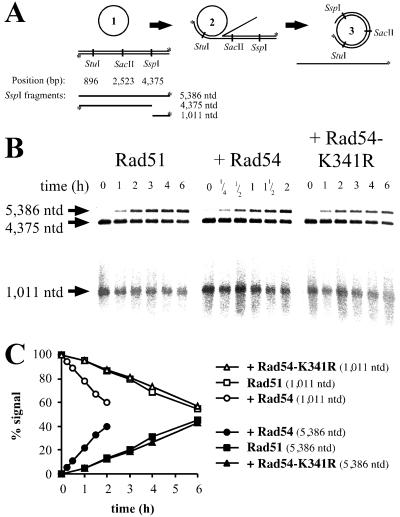
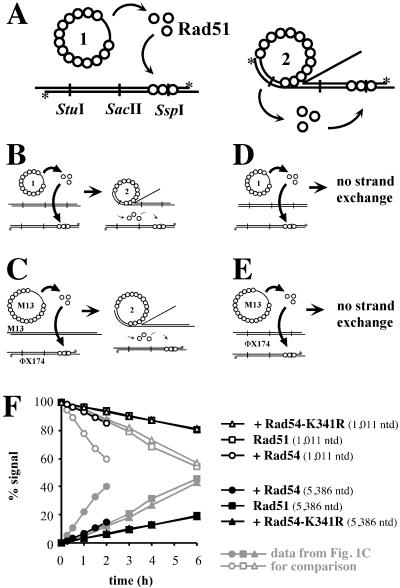
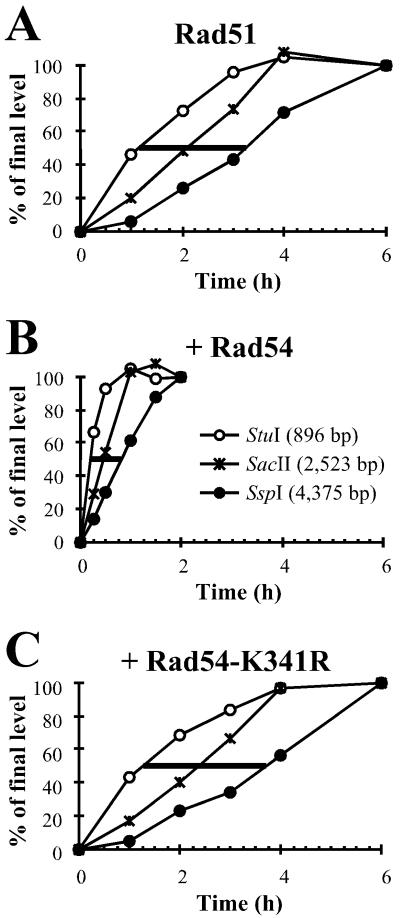
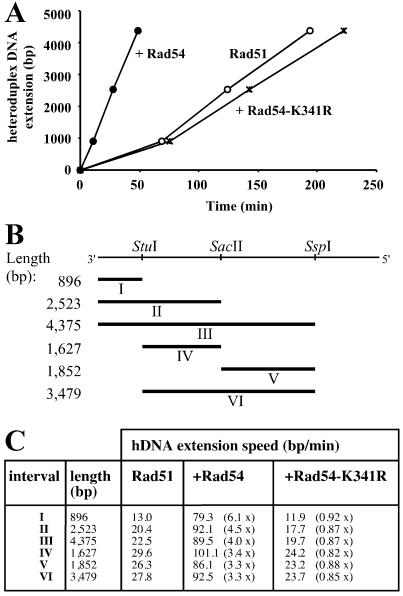
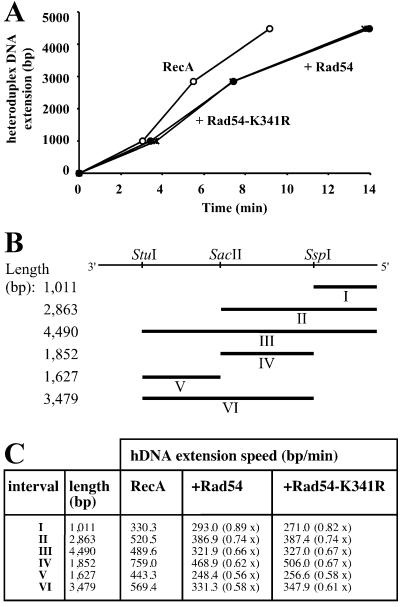
Rad54 and Rad51 are important proteins for the repair of double-stranded DNA breaks by homologous recombination in eukaryotes. As previously shown, Rad51 protein forms nucleoprotein filaments on single-stranded DNA, and Rad54 protein directly interacts with such filaments to enhance synapsis, the homologous pairing with a double-stranded DNA partner. Here we demonstrate that Saccharomyces cerevisiae Rad54 protein has an additional role in the postsynaptic phase of DNA strand exchange by stimulating heteroduplex DNA extension of established joint molecules in Rad51/Rpa-mediated DNA strand exchange. This function depended on the ATPase activity of Rad54 protein and on specific protein:protein interactions between the yeast Rad54 and Rad51 proteins.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
