

Simian virus 40 Vp1 DNA-binding domain is functionally separable from the overlapping nuclear localization signal and is required for effective virion formation and full viability
- PMID: 11462004
- PMCID: PMC114967
- DOI: 10.1128/JVI.75.16.7321-7329.2001
Simian virus 40 Vp1 DNA-binding domain is functionally separable from the overlapping nuclear localization signal and is required for effective virion formation and full viability
Abstract
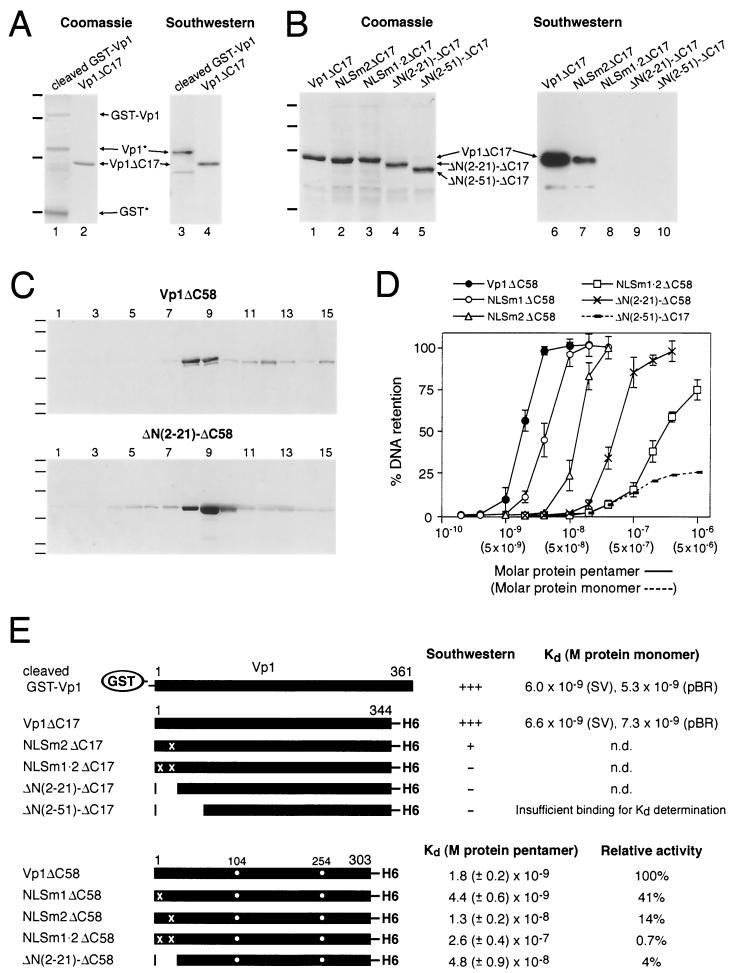
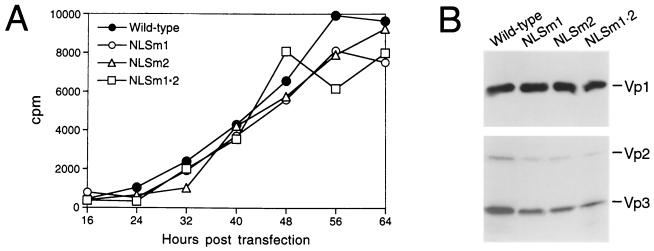
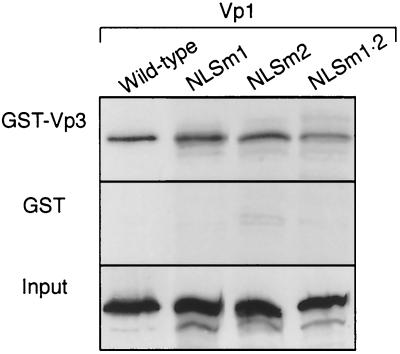
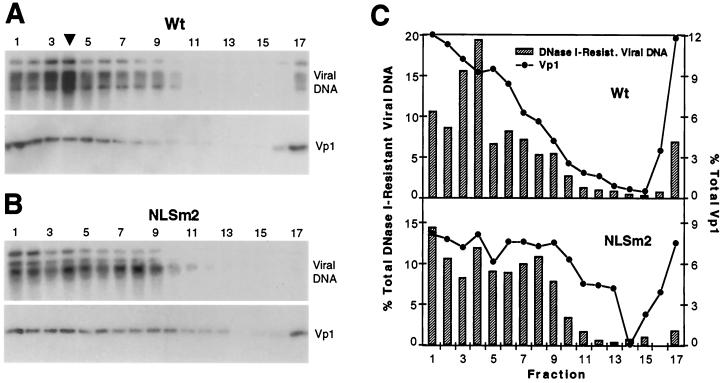
A DNA-binding domain (DBD) was identified on simian virus 40 (SV40) major capsid protein Vp1, and the domain's function in the SV40 life cycle was examined. The DBD was mapped by assaying various recombinant Vp1 proteins for DNA binding in vitro. The carboxy-terminal 58-residue truncated Vp1DeltaC58 pentamer bound DNA with a K(d) of 1.8 x 10(-9) M in terms of the protein pentamer, while full-length Vp1 and carboxy-terminal-17-truncated Vp1DeltaC17 had comparable apparent K(d)s of 5.3 x 10(-9) to 7.3 x 10(-9) M in terms of the protein monomers. Previously identified on Vp1 was a nuclear localization signal (NLS) consisting of two N-terminal basic clusters, NLS1 (4-KRK-6) and NLS2 (15-KKPK-18). Vp1DeltaC58 pentamers harboring multiple-point mutations in NLS1 (NLSm1), NLS2 (NLSm2), or both basic clusters (NLSm1. 2) had progressively decreased DNA-binding activity, down to 0.7% of the Vp1DeltaC58 level for NLSm1. 2 Vp1. These data, along with those of N-terminally truncated proteins, placed the DBD in overlap with the bipartite NLS. The role of the Vp1 DBD during infection was investigated by taking advantage of NLS phenotypic complementation (N. Ishii, A. Nakanishi, M. Yamada, M. H. Macalalad, and H. Kasamatsu, J. Virol. 68:8209-8216, 1994), in which an NLS-defective Vp1 could localize to the nucleus in the presence of wild-type minor capsid proteins Vp2 and Vp3. This approach made it possible to dissect the role of the bifunctional Vp1 NLS-DBD in virion assembly in the nucleus. Mutants of the viable nonoverlapping SV40 (NO-SV40) DNA NLSm1, NLSm2, and NLSm1. 2 replicated normally following transfection into host cells and produced capsid proteins at normal levels. All mutant Vp1s were able to interact with Vp3 in vitro. The mutants NLSm1 and NLSm1. 2 were nonviable, and the mutant Vp1s unexpectedly failed to localize to the nucleus though Vp2 and Vp3 did, suggesting that the mutated NLS1 acted as a dominant signal for the cytoplasmic localization of Vp1. Mutant NLSm2, for which the mutant Vp1's nuclear localization defect was complemented by Vp2 and Vp3, displayed a 5,000-fold reduced viability. Analysis of NLSm2 DNA-transfected cell lysate revealed a 10-fold reduction in the level of DNase I-protected viral DNA, and yet virion-like particles were found among the DNase I-resistant material. Collective results support a role for Vp1 NLS2-DBD2 in the assembly of virion particles. The results also suggest that this determinant can function in the infection of new cells.
Figures


Similar articles
-
Minor capsid proteins of simian virus 40 are dispensable for nucleocapsid assembly and cell entry but are required for nuclear entry of the viral genome.J Virol. 2007 Apr;81(8):3778-85. doi: 10.1128/JVI.02664-06. Epub 2007 Jan 31. J Virol. 2007. PMID: 17267496 Free PMC article.
-
Role of simian virus 40 Vp1 cysteines in virion infectivity.J Virol. 2000 Dec;74(23):11388-93. doi: 10.1128/jvi.74.23.11388-11393.2000. J Virol. 2000. PMID: 11070039 Free PMC article.
-
Essential role of the Vp2 and Vp3 DNA-binding domain in simian virus 40 morphogenesis.J Virol. 1995 Feb;69(2):1115-21. doi: 10.1128/JVI.69.2.1115-1121.1995. J Virol. 1995. PMID: 7815491 Free PMC article.
-
Simian virus 40 chromatin interaction with the capsid proteins.J Biomol Struct Dyn. 1983 Dec;1(3):689-704. doi: 10.1080/07391102.1983.10507475. J Biomol Struct Dyn. 1983. PMID: 6101085 Review.
-
Host range analysis of simian virus 40, BK virus and chimaeric SV40/BKV: relative expression of large T-antigen and Vp1 in infected and transformed cells.Dev Biol Stand. 1998;94:191-205. Dev Biol Stand. 1998. PMID: 9776240 Review.
Cited by
-
Minor capsid proteins of simian virus 40 are dispensable for nucleocapsid assembly and cell entry but are required for nuclear entry of the viral genome.J Virol. 2007 Apr;81(8):3778-85. doi: 10.1128/JVI.02664-06. Epub 2007 Jan 31. J Virol. 2007. PMID: 17267496 Free PMC article.
-
Identification of amino acid residues within simian virus 40 capsid proteins Vp1, Vp2, and Vp3 that are required for their interaction and for viral infection.J Virol. 2006 Sep;80(18):8891-8. doi: 10.1128/JVI.00781-06. J Virol. 2006. PMID: 16940501 Free PMC article.
-
Possible role for cellular karyopherins in regulating polyomavirus and papillomavirus capsid assembly.J Virol. 2008 Oct;82(20):9848-57. doi: 10.1128/JVI.01221-08. Epub 2008 Aug 13. J Virol. 2008. PMID: 18701594 Free PMC article.
-
Molecular dissection of nuclear entry-competent SV40 during infection.Virus Res. 2007 Mar;124(1-2):226-30. doi: 10.1016/j.virusres.2006.10.001. Epub 2006 Nov 16. Virus Res. 2007. PMID: 17112617 Free PMC article.
-
Assembly and Stability of Simian Virus 40 Polymorphs.ACS Nano. 2020 Apr 28;14(4):4430-4443. doi: 10.1021/acsnano.9b10004. Epub 2020 Apr 2. ACS Nano. 2020. PMID: 32208635 Free PMC article.
References
-
- Blasquez V, Beecher S, Bina M. Simian virus 40 morphogenetic pathway. J Biol Chem. 1983;258:8477–8484. - PubMed
-
- Clever J, Kasamatsu H. Simian virus 40 Vp2/3 small structural proteins harbor their own nuclear transport signal. Virology. 1991;181:78–90. - PubMed
-
- Clever J, Dean D A, Kasamatsu H. Identification of a DNA-binding domain in simian virus 40 capsid proteins Vp2 and Vp3. J Biol Chem. 1993;268:20877–20883. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
