

Influenza B and C virus NEP (NS2) proteins possess nuclear export activities
- PMID: 11462009
- PMCID: PMC114972
- DOI: 10.1128/JVI.75.16.7375-7383.2001
Influenza B and C virus NEP (NS2) proteins possess nuclear export activities
Abstract
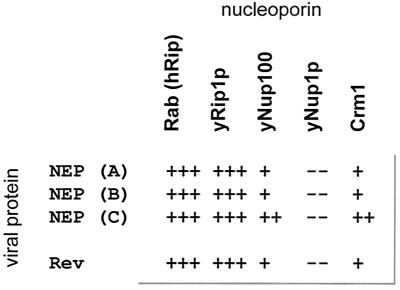
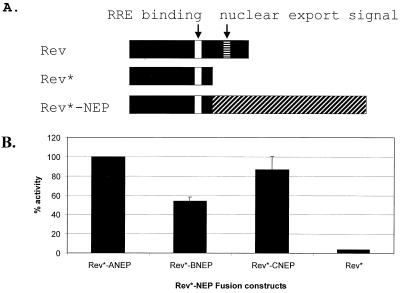
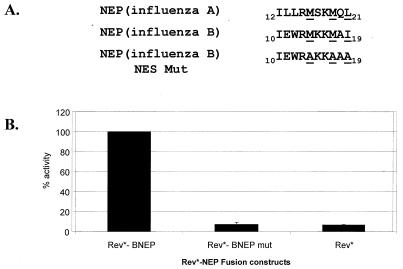
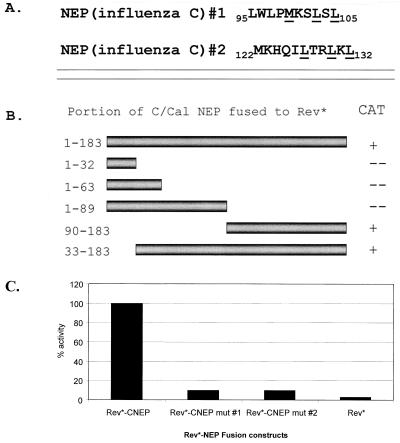
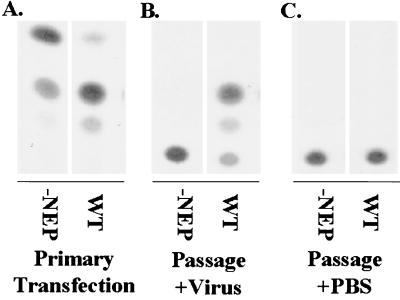
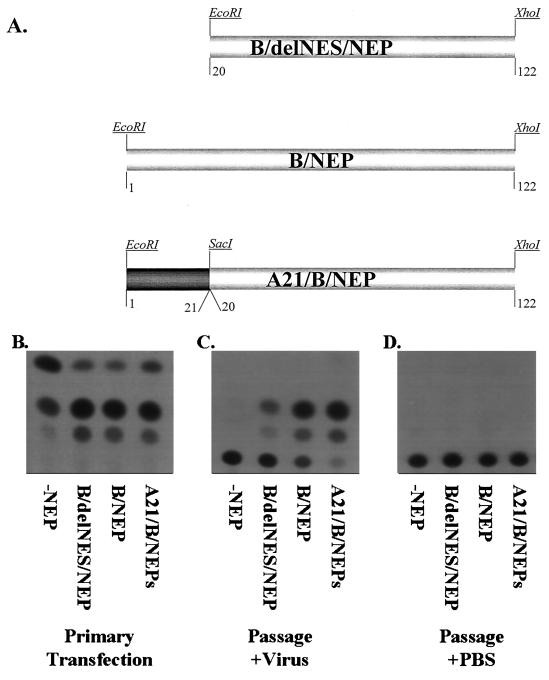
Nucleocytoplasmic transport of viral ribonucleoproteins (vRNPs) is an essential aspect of the replication cycle for influenza A, B, and C viruses. These viruses replicate and transcribe their genomes in the nuclei of infected cells. During the late stages of infection, vRNPs must be exported from the nucleus to the cytoplasm prior to transport to viral assembly sites on the cellular plasma membrane. Previously, we demonstrated that the influenza A virus nuclear export protein (NEP, formerly referred to as the NS2 protein) mediates the export of vRNPs. In this report, we suggest that for influenza B and C viruses the nuclear export function is also performed by the orthologous NEP proteins (formerly referred to as the NS2 protein). The influenza virus B and C NEP proteins interact in the yeast two-hybrid assay with a subset of nucleoporins and with the Crm1 nuclear export factor and can functionally replace the effector domain from the human immunodeficiency virus type 1 Rev protein. We established a plasmid transfection system for the generation of virus-like particles (VLPs) in which a functional viral RNA-like chloramphenicol acetyltransferase (CAT) gene is delivered to a new cell. VLPs generated in the absence of the influenza B virus NEP protein were unable to transfer the viral RNA-like CAT gene to a new cell. From these data, we suggest that the nuclear export of the influenza B and C vRNPs are mediated through interaction between NEP proteins and the cellular nucleocytoplasmic export machinery.
Figures
Similar articles
-
Intracellular localization of influenza C virus NS2 protein (NEP) in infected cells and its incorporation into virions.Arch Virol. 2009;154(2):235-43. doi: 10.1007/s00705-008-0292-6. Epub 2009 Jan 8. Arch Virol. 2009. PMID: 19130168
-
Generation of influenza A virus NS2 (NEP) mutants with an altered nuclear export signal sequence.J Virol. 2004 Sep;78(18):10149-55. doi: 10.1128/JVI.78.18.10149-10155.2004. J Virol. 2004. PMID: 15331747 Free PMC article.
-
Crucial role of the influenza virus NS2 (NEP) C-terminal domain in M1 binding and nuclear export of vRNP.FEBS Lett. 2011 Jan 3;585(1):41-6. doi: 10.1016/j.febslet.2010.11.017. Epub 2010 Nov 16. FEBS Lett. 2011. PMID: 21081124
-
Nuclear functions of the influenza A and B viruses NS1 proteins: do they play a role in viral mRNA export?Vaccine. 2009 Oct 23;27(45):6312-6. doi: 10.1016/j.vaccine.2009.01.015. Vaccine. 2009. PMID: 19840666 Review.
-
Nuclear traffic of influenza virus proteins and ribonucleoprotein complexes.Virus Res. 2007 Mar;124(1-2):12-21. doi: 10.1016/j.virusres.2006.09.013. Epub 2006 Nov 1. Virus Res. 2007. PMID: 17081640 Review.
Cited by
-
Role of the CM2 protein in the influenza C virus replication cycle.J Virol. 2011 Feb;85(3):1322-9. doi: 10.1128/JVI.01367-10. Epub 2010 Nov 24. J Virol. 2011. PMID: 21106743 Free PMC article.
-
Influenza A in Bovine Species: A Narrative Literature Review.Viruses. 2019 Jun 17;11(6):561. doi: 10.3390/v11060561. Viruses. 2019. PMID: 31213032 Free PMC article. Review.
-
Phosphorylation of S-S-S Motif in Nuclear Export Protein (NEP) Plays a Critical Role in Viral Ribonucleoprotein (vRNP) Nuclear Export of Influenza A and B Viruses.Adv Sci (Weinh). 2025 Jan;12(2):e2309477. doi: 10.1002/advs.202309477. Epub 2024 Nov 22. Adv Sci (Weinh). 2025. PMID: 39575547 Free PMC article.
-
Peptide-mediated interference with influenza A virus polymerase.J Virol. 2007 Jul;81(14):7801-4. doi: 10.1128/JVI.00724-07. Epub 2007 May 9. J Virol. 2007. PMID: 17494067 Free PMC article.
-
Establishment of a Reverse Genetics System for Influenza D Virus.J Virol. 2020 May 4;94(10):e01767-19. doi: 10.1128/JVI.01767-19. Print 2020 May 4. J Virol. 2020. PMID: 32102883 Free PMC article.
References
-
- Alamgir A S, Matsuzaki Y, Hongo S, Tsuchiya E, Sugawara K, Muraki Y, Nakamura K. Phylogenetic analysis of influenza C virus nonstructural (NS) protein genes and identification of the NS2 protein. J Gen Virol. 2000;81:1933–1940. - PubMed
-
- Basler C, Palese P. Influenza viruses. In: Creighton T, editor. Encyclopedia of molecular medicine. New York, N.Y: John Wiley and Sons; 2001.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
