

Reconstitution of a functional duck hepatitis B virus replication initiation complex from separate reverse transcriptase domains expressed in Escherichia coli
- PMID: 11462013
- PMCID: PMC114976
- DOI: 10.1128/JVI.75.16.7410-7419.2001
Reconstitution of a functional duck hepatitis B virus replication initiation complex from separate reverse transcriptase domains expressed in Escherichia coli
Abstract
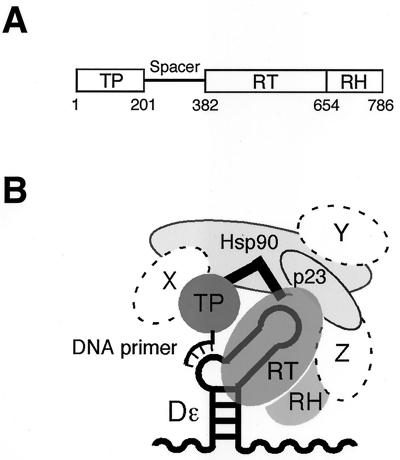
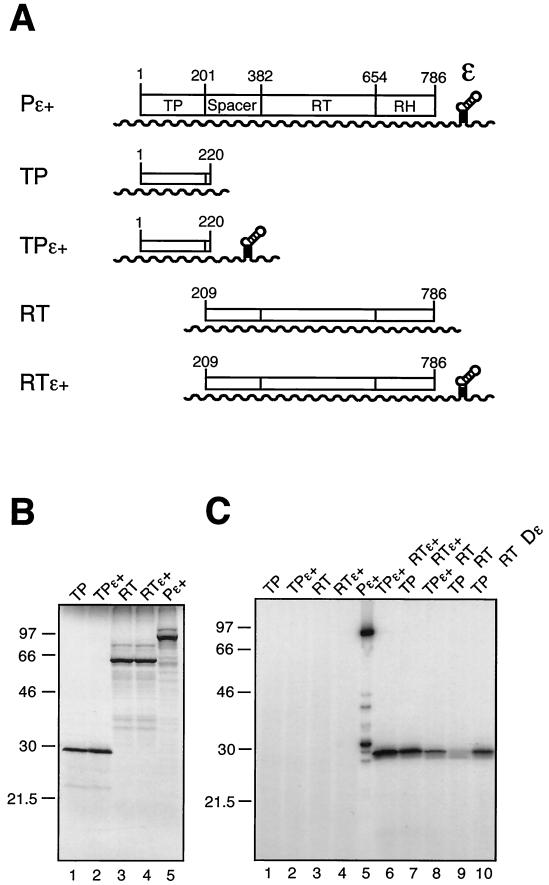
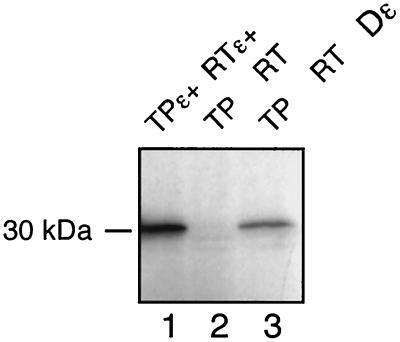
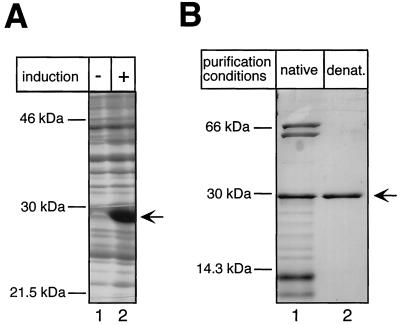
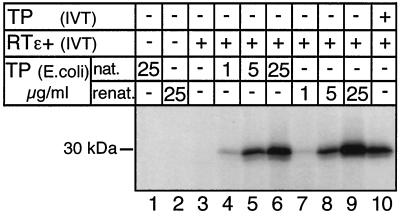
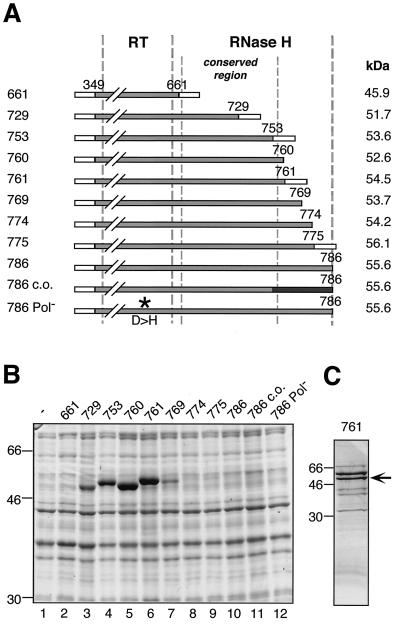
Hepatitis B viruses replicate through reverse transcription of an RNA intermediate, the pregenomic RNA (pgRNA). Replication is initiated de novo and requires formation of a ribonucleoprotein complex comprising the viral reverse transcriptase (P protein), an RNA stem-loop structure (epsilon) on the pgRNA, and cellular proteins, including the heat shock protein Hsp90, the cochaperone p23, and additional, as yet unknown, factors. Functional complexes catalyze the synthesis of a short DNA primer that is templated by epsilon and covalently linked to the terminal protein (TP) domain of P protein. Currently, the only system for generating such complexes in the test tube is in vitro translation of duck hepatitis B virus (DHBV) P protein in rabbit reticulocyte lysate (RRL), which also provides the necessary factors. However, its limited translation capacity precludes a closer analysis of the complex. To overcome this restriction we sought to produce larger amounts of DHBV P protein by expression in Escherichia coli, followed by complex reconstitution in RRL. Because previous attempts to generate full-length P protein in bacteria have failed we investigated whether separate expression of the TP and reverse transcriptase-RNase H (RT-RH) domains would allow higher yields and whether these domains could trans complement each other. Indeed, TP and, after minor C-terminal modifications, also RT-RH could be expressed in substantial amounts, and when added to RRL, they were capable of epsilon-dependent DNA primer synthesis, demonstrating posttranslational activation. This reconstitution system should pave the way for a detailed understanding of the unique hepadnaviral replication initiation mechanism.
Figures


References
-
- Beck J, Bartos H, Nassal M. Experimental confirmation of a hepatitis B virus (HBV) epsilon like bulge and loop structure in avian HBV RNA encapsidation signals. Virology. 1997;227:500–504. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
