

Domain interactions in the yeast ATP binding cassette transporter Ycf1p: intragenic suppressor analysis of mutations in the nucleotide binding domains
- PMID: 11466279
- PMCID: PMC99530
- DOI: 10.1128/JB.183.16.4761-4770.2001
Domain interactions in the yeast ATP binding cassette transporter Ycf1p: intragenic suppressor analysis of mutations in the nucleotide binding domains
Abstract
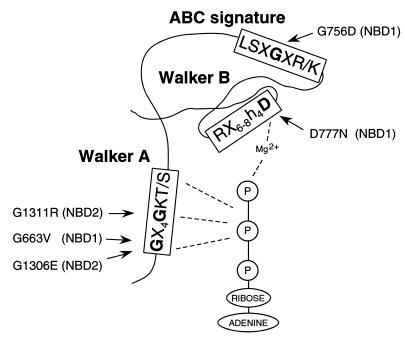
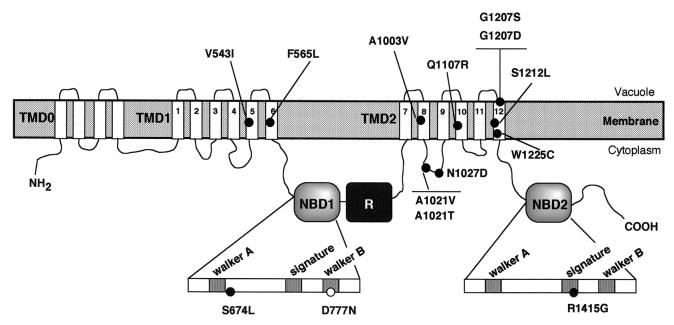
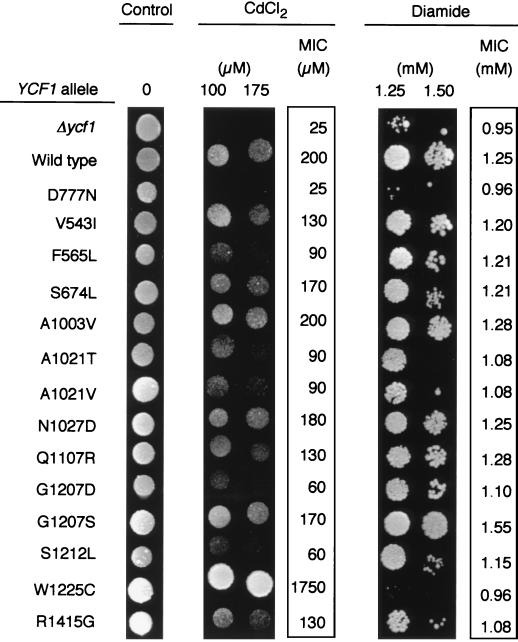
The yeast cadmium factor (Ycf1p) is a vacuolar ATP binding cassette (ABC) transporter required for heavy metal and drug detoxification. Cluster analysis shows that Ycf1p is strongly related to the human multidrug-associated protein (MRP1) and cystic fibrosis transmembrane conductance regulator and therefore may serve as an excellent model for the study of eukaryotic ABC transporter structure and function. Identifying intramolecular interactions in these transporters may help to elucidate energy transfer mechanisms during transport. To identify regions in Ycf1p that may interact to couple ATPase activity to substrate binding and/or movement across the membrane, we sought intragenic suppressors of ycf1 mutations that affect highly conserved residues presumably involved in ATP binding and/or hydrolysis. Thirteen intragenic second-site suppressors were identified for the D777N mutation which affects the invariant Asp residue in the Walker B motif of the first nucleotide binding domain (NBD1). Two of the suppressor mutations (V543I and F565L) are located in the first transmembrane domain (TMD1), nine (A1003V, A1021T, A1021V, N1027D, Q1107R, G1207D, G1207S, S1212L, and W1225C) are found within TMD2, one (S674L) is in NBD1, and another one (R1415G) is in NBD2, indicating either physical proximity or functional interactions between NBD1 and the other three domains. The original D777N mutant protein exhibits a strong defect in the apparent affinity for ATP and V(max) of transport. The phenotypic characterization of the suppressor mutants shows that suppression does not result from restoring these alterations but rather from a change in substrate specificity. We discuss the possible involvement of Asp777 in coupling ATPase activity to substrate binding and/or transport across the membrane.
Figures


References
-
- Anderson M P, Gregory R J, Thompson S, Souza D W, Paul S, Mulligan R C, Smith A E, Welsh M J. Demonstration that CFTR is a chloride channel by alteration of its anion selectivity. Science. 1991;253:202–205. - PubMed
-
- Armstrong S, Tabernero L, Zhang H, Hermodson M, Stauffacher C. Powering the ABC transporter: the 2.5 Å crystallographic structure of the ABC domain of RBSA. Pediatr Pulmonol. 1998;26:91–92.
-
- Blake M S, Johnston K H, Russell-Jones G H, Gotschlich E C. A rapid, sensitive method for detection of alkaline phosphatase-conjugated anti-antibody on Western blots. Anal Biochem. 1984;136:175–179. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
