

Behavioral properties of the trigeminal somatosensory system in rats performing whisker-dependent tactile discriminations
- PMID: 11466447
- PMCID: PMC6762640
- DOI: 10.1523/JNEUROSCI.21-15-05752.2001
Behavioral properties of the trigeminal somatosensory system in rats performing whisker-dependent tactile discriminations
Abstract
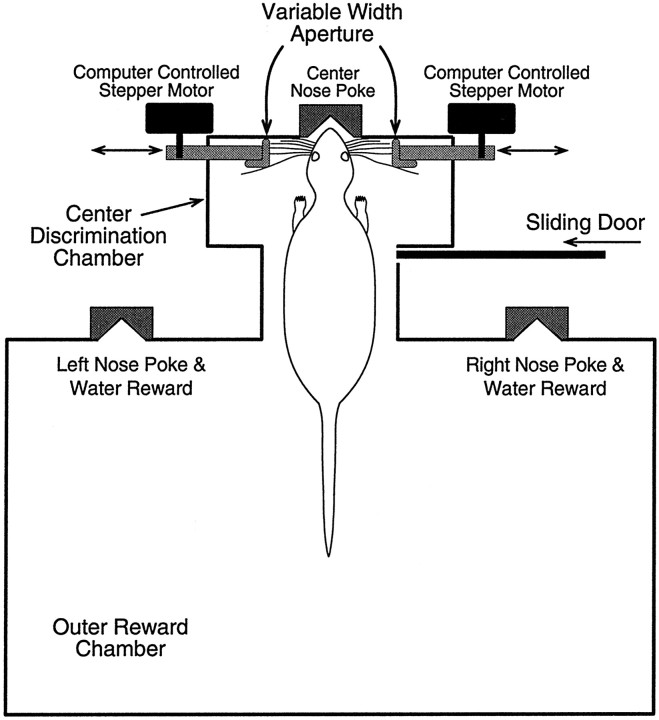
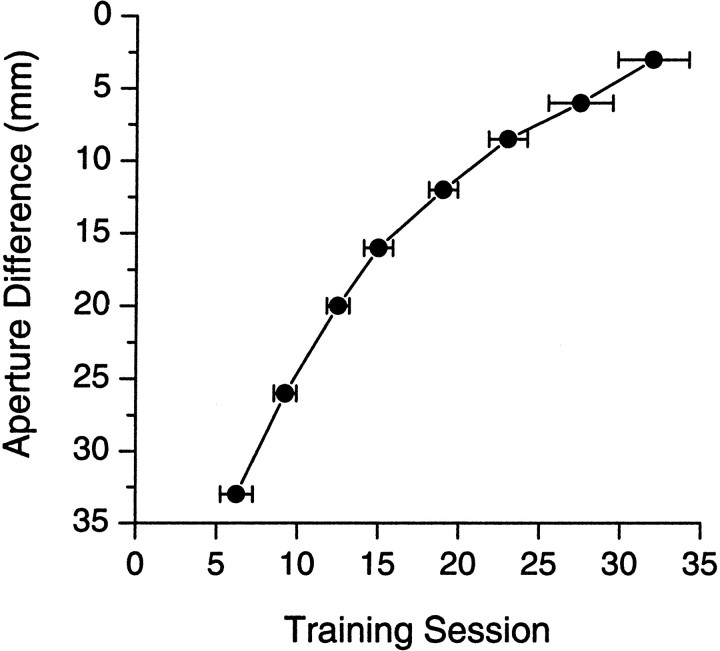
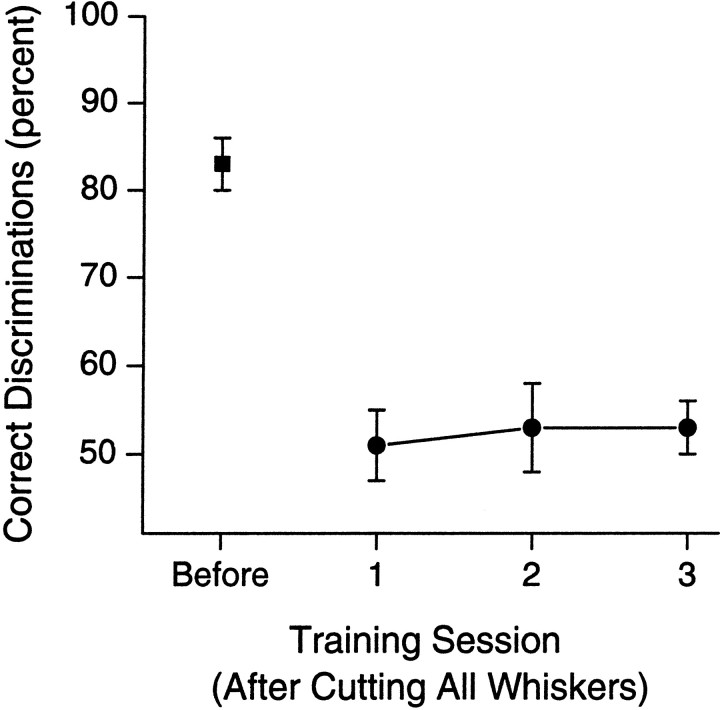
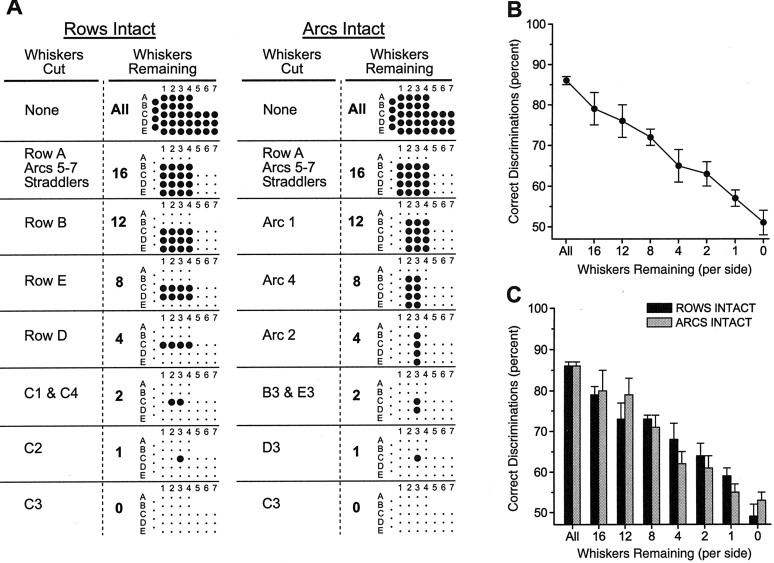
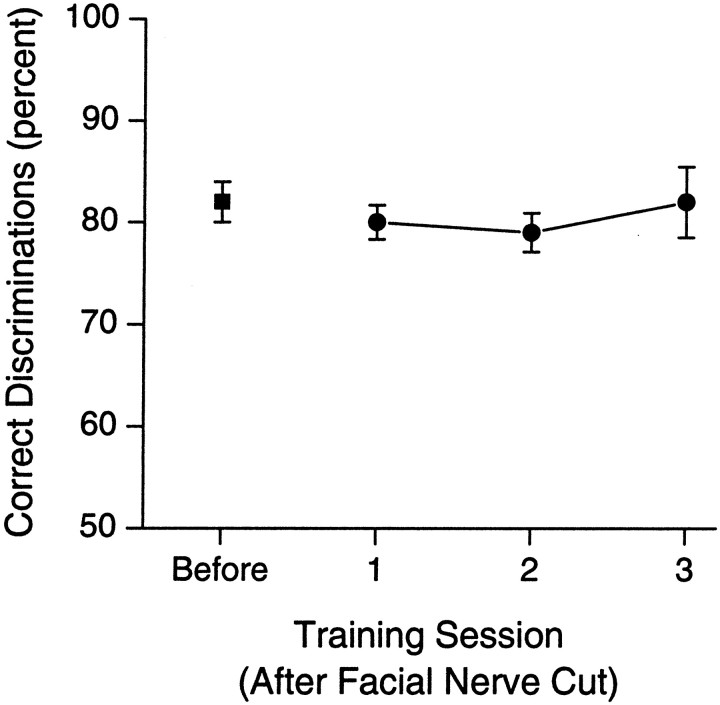
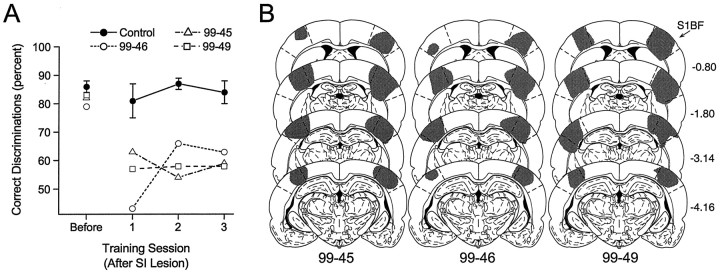
To address several fundamental questions regarding how multiwhisker tactile stimuli are integrated and processed by the trigeminal somatosensory system, a novel behavioral task was developed that required rats to discriminate the width of either a wide or narrow aperture using only their large mystacial vibrissae. Rats quickly acquired this task and could accurately discriminate between apertures of very similar width. Accurate discriminations required a large number of intact facial whiskers. Systematic removal of individual whiskers caused a decrease in performance that was directly proportional to the number of whiskers removed, indicating that tactile information from multiple whiskers is integrated as rats gauge aperture width. In different groups of rats, different sets of whiskers were removed in patterns that preferentially left whisker rows or whisker arcs intact. These different whisker removals caused similar decreases in performance, indicating that individual whiskers within the vibrissal array are functionally equivalent during performance of this task. Lesions of the barrel cortex abolished the ability of rats to discriminate, demonstrating that this region is critically involved in this tactile behavior. Interestingly, sectioning the facial nerve, which abolished whisker movements, did not affect the ability to perform accurate discriminations, indicating that active whisker movements are not necessary for accurate performance of the task. Collectively, these results indicate that the trigeminal somatosensory system forms internal representations of external stimuli (in this case, aperture width) by integrating tactile input from many functionally equivalent facial whiskers and that the vibrissal array can function as a fine-grained distance detector without active whisker movements.
Figures

References
-
- Armstrong-James M, Fox K. Spatiotemporal convergence and divergence in the rat S1 “barrel” cortex. J Comp Neurol. 1987;263:265–281. - PubMed
-
- Arvidsson J. Somatotopic organization of vibrissae afferents in the trigeminal sensory nuclei of the rat studied by transganglionic transport of HRP. J Comp Neurol. 1982;211:84–92. - PubMed
-
- Belford GR, Killackey HP. Vibrissae representation in subcortical trigeminal centers of the neonatal rat. J Comp Neurol. 1979;183:305–321. - PubMed
-
- Brecht M, Preilowski B, Merzenich MM. Functional architecture of the mystacial vibrissae. Behav Brain Res. 1997;84:81–97. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources