

Regulation of limb patterning by extracellular microfibrils
- PMID: 11470817
- PMCID: PMC2150751
- DOI: 10.1083/jcb.200105046
Regulation of limb patterning by extracellular microfibrils
Abstract
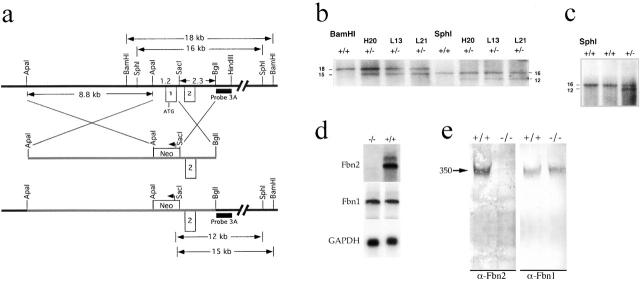
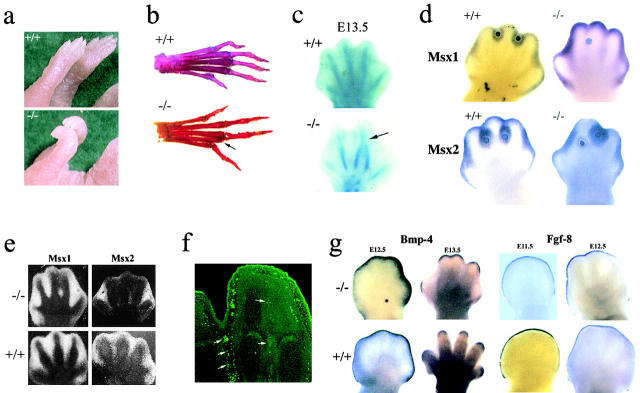
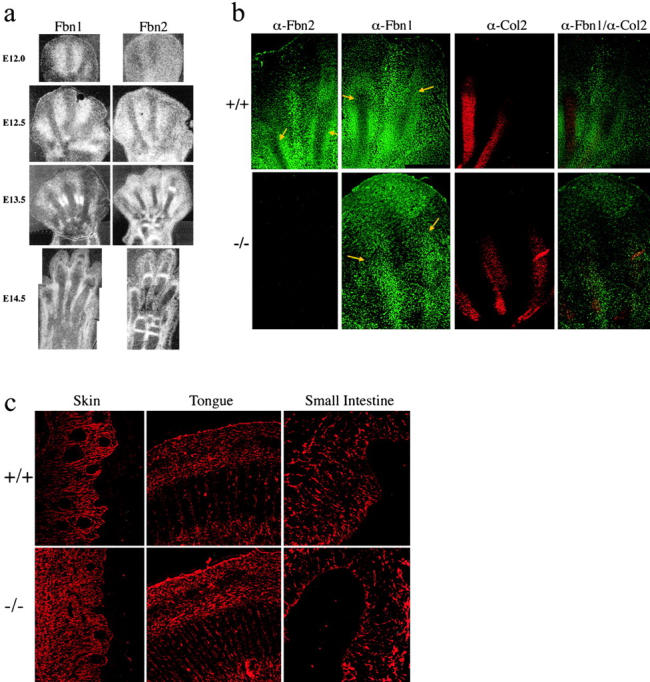
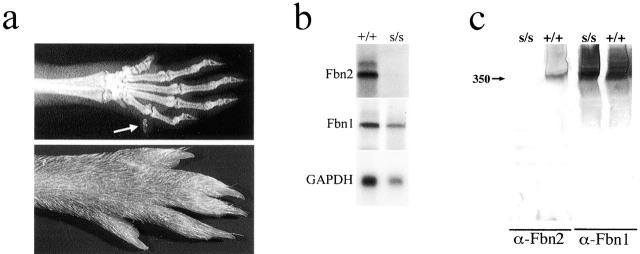
To elucidate the contribution of the extracellular microfibril-elastic fiber network to vertebrate organogenesis, we generated fibrillin 2 (Fbn2)-null mice by gene targeting and identified a limb-patterning defect in the form of bilateral syndactyly. Digit fusion involves both soft and hard tissues, and is associated with reduced apoptosis at affected sites. Two lines of evidence suggest that syndactily is primarily due to defective mesenchyme differentiation, rather than reduced apoptosis of interdigital tissue. First, fusion occurs before appearance of interdigital cell death; second, interdigital tissues having incomplete separation fail to respond to apoptotic clues from implanted BMP-4 beads. Syndactyly is associated with a disorganized matrix, but with normal BMP gene expression. On the other hand, mice double heterozygous for null Fbn2 and Bmp7 alleles display the combined digit phenotype of both nullizygotes. Together, these results imply functional interaction between Fbn2-rich microfibrils and BMP-7 signaling. As such, they uncover an unexpected relationship between the insoluble matrix and soluble factors during limb patterning. We also demonstrate that the Fbn2- null mutation is allelic to the recessive shaker-with-syndactyly (sy) locus on chromosome 18.
Figures
References
-
- Chen, Y., and X. Zhao. 1998. Shaping limbs by apoptosis. J. Exp. Zool. 282:691–702. - PubMed
-
- Christian, J.L. 2000. BMP, Wnt and Hedgehog signals: how far can they go? Curr. Opin. Cell Biol. 12:244–249. - PubMed
-
- Dahna, R.D., and J.F. Fallon. 2000. Interdigital regulation of digit identity and homeotic transformation by modulated BMP signaling. Science. 289:438–441. - PubMed
-
- Dudley, A.T., Lyons K.M., and Robertson E.J. 1995. A requirement for bone morphogenetic protein-7 during development of the mammalian kidney and eye. Genes Dev. 9:2795–2807. - PubMed
-
- Flaumenhaft, R., and D.B. Rifkin. 1991. Extracellular matrix regulation of growth factor and protease activity. Curr. Opin. Cell Biol. 3:817–823. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
