

Molecular architecture of the voltage-dependent Na channel: functional evidence for alpha helices in the pore
- PMID: 11479344
- PMCID: PMC2233827
- DOI: 10.1085/jgp.118.2.171
Molecular architecture of the voltage-dependent Na channel: functional evidence for alpha helices in the pore
Abstract
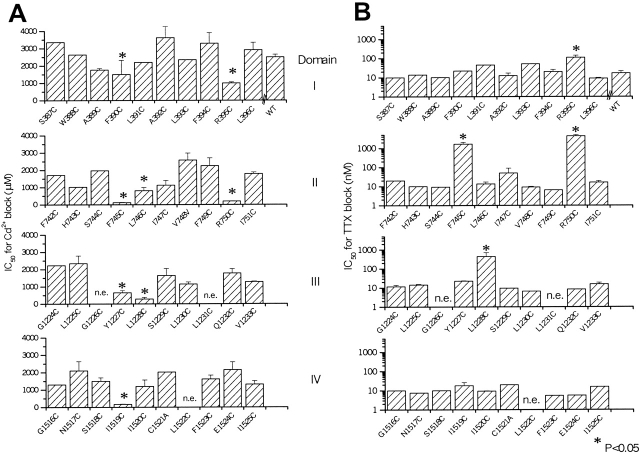
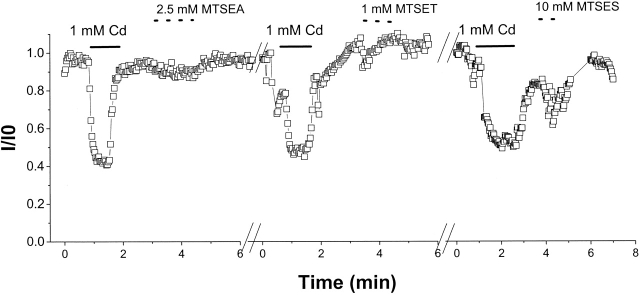
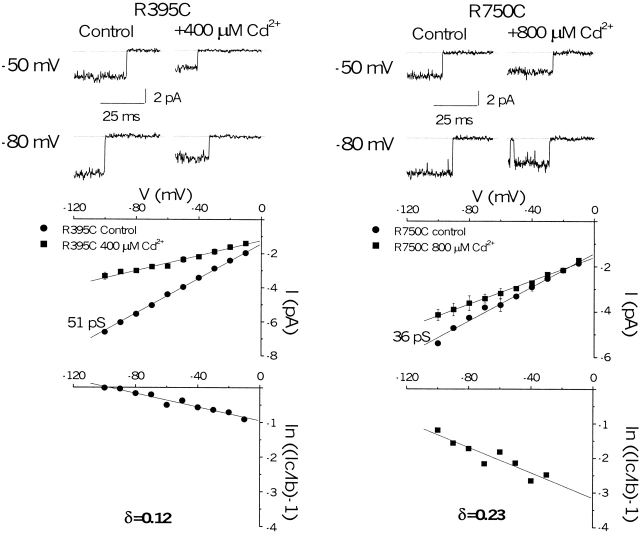
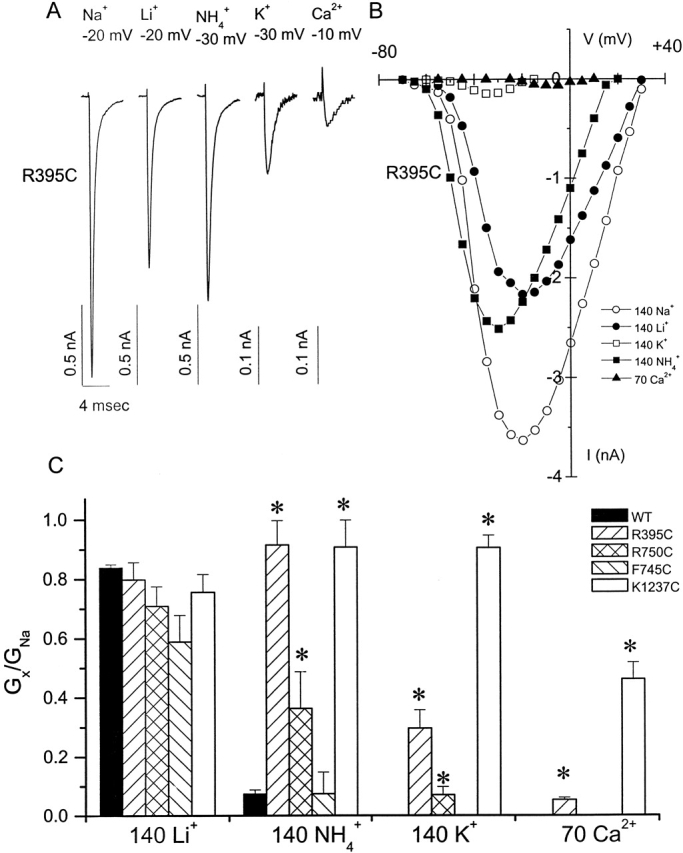
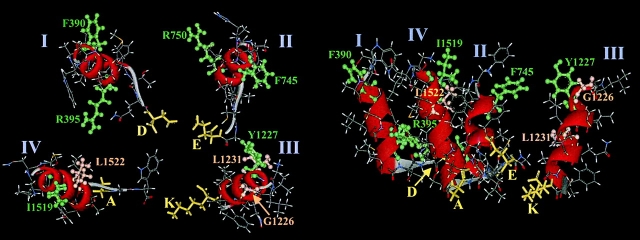
The permeation pathway of the Na channel is formed by asymmetric loops (P segments) contributed by each of the four domains of the protein. In contrast to the analogous region of K channels, previously we (Yamagishi, T., M. Janecki, E. Marban, and G. Tomaselli. 1997. Biophys. J. 73:195-204) have shown that the P segments do not span the selectivity region, that is, they are accessible only from the extracellular surface. The portion of the P-segment NH(2)-terminal to the selectivity region is referred to as SS1. To explore further the topology and functional role of the SS1 region, 40 amino acids NH(2)-terminal to the selectivity ring (10 in each of the P segments) of the rat skeletal muscle Na channel were substituted by cysteine and expressed in tsA-201 cells. Selected mutants in each domain could be blocked with high affinity by externally applied Cd(2)+ and were resistant to tetrodotoxin as compared with the wild-type channel. None of the externally applied sulfhydryl-specific methanethiosulfonate reagents modified the current through any of the mutant channels. Both R395C and R750C altered ionic selectivity, producing significant increases in K(+) and NH(4)(+) currents. The pattern of side chain accessibility is consistent with a pore helix like that observed in the crystal structure of the bacterial K channel, KcsA. Structure prediction of the Na channel using the program PHDhtm suggests an alpha helix in the SS1 region of each domain channel. We conclude that each of the P segments undergoes a hairpin turn in the permeation pathway, such that amino acids on both sides of the putative selectivity filter line the outer mouth of the pore. Evolutionary conservation of the pore helix motif from bacterial K channels to mammalian Na channels identifies this structure as a critical feature in the architecture of ion selective pores.
Figures


Similar articles
-
A tryptophan residue (W736) in the amino-terminus of the P-segment of domain II is involved in pore formation in Na(v)1.4 voltage-gated sodium channels.Pflugers Arch. 2002 Oct;445(1):18-24. doi: 10.1007/s00424-002-0887-9. Epub 2002 Aug 14. Pflugers Arch. 2002. PMID: 12397382
-
Topology of the P segments in the sodium channel pore revealed by cysteine mutagenesis.Biophys J. 1997 Jul;73(1):195-204. doi: 10.1016/S0006-3495(97)78060-3. Biophys J. 1997. PMID: 9199784 Free PMC article.
-
KcsA crystal structure as framework for a molecular model of the Na(+) channel pore.Biochemistry. 2000 Jul 18;39(28):8161-70. doi: 10.1021/bi000486w. Biochemistry. 2000. PMID: 10889022
-
Molecular pore structure of voltage-gated sodium and calcium channels.Braz J Med Biol Res. 1994 Dec;27(12):2781-802. Braz J Med Biol Res. 1994. PMID: 7550000 Review.
-
Molecular properties of brain sodium channels: an important target for anticonvulsant drugs.Adv Neurol. 1999;79:441-56. Adv Neurol. 1999. PMID: 10514834 Review.
Cited by
-
Models of the structure and gating mechanisms of the pore domain of the NaChBac ion channel.Biophys J. 2008 Oct;95(8):3650-62. doi: 10.1529/biophysj.108.135327. Epub 2008 Jul 18. Biophys J. 2008. PMID: 18641075 Free PMC article.
-
Three ENU-induced neurological mutations in the pore loop of sodium channel Scn8a (Na(v)1.6) and a genetically linked retinal mutation, rd13.Mamm Genome. 2004 May;15(5):344-51. doi: 10.1007/s00335-004-2332-1. Mamm Genome. 2004. PMID: 15170223
-
Sodium channels: ionic model of slow inactivation and state-dependent drug binding.Biophys J. 2007 Sep 1;93(5):1557-70. doi: 10.1529/biophysj.106.100248. Epub 2007 May 11. Biophys J. 2007. PMID: 17496040 Free PMC article.
-
Congenital insensitivity to pain: novel SCN9A missense and in-frame deletion mutations.Hum Mutat. 2010 Sep;31(9):E1670-86. doi: 10.1002/humu.21325. Hum Mutat. 2010. PMID: 20635406 Free PMC article.
-
Constraint shapes convergence in tetrodotoxin-resistant sodium channels of snakes.Proc Natl Acad Sci U S A. 2012 Mar 20;109(12):4556-61. doi: 10.1073/pnas.1113468109. Epub 2012 Mar 5. Proc Natl Acad Sci U S A. 2012. PMID: 22392995 Free PMC article.
References
-
- Backx P.H., Yue D.T., Lawrence J.H., Marban E., Tomaselli G.F. Molecular localization of an ion-binding site within the pore of mammalian sodium channels. Science. 1992;257:248–251. - PubMed
-
- Catterall W.A. Structure and function of voltage-gated ion channels. Annu. Rev. Biochem. 1995;64:493–531. - PubMed
-
- Chang N.S., French R.J., Lipkind G.M., Fozzard H.A., Dudley S., Jr. Predominant interactions between mu-conotoxin Arg-13 and the skeletal muscle Na+ channel localized by mutant cycle analysis. Biochemistry. 1998;37:4407–4419. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
