

A genomic island in Pseudomonas aeruginosa carries the determinants of flagellin glycosylation
- PMID: 11481492
- PMCID: PMC55422
- DOI: 10.1073/pnas.161249198
A genomic island in Pseudomonas aeruginosa carries the determinants of flagellin glycosylation
Abstract
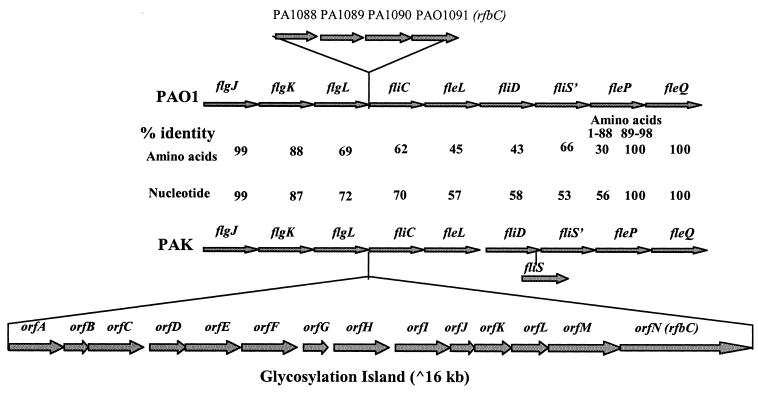
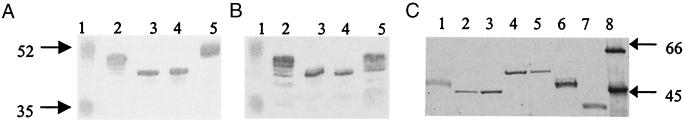
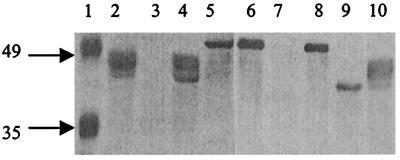
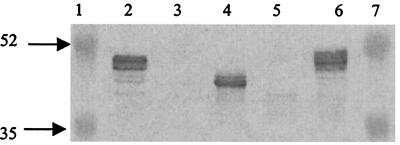
Protein glycosylation has been long recognized as an important posttranslational modification process in eukaryotic cells. Glycoproteins, predominantly secreted or surface localized, have also been identified in bacteria. We have identified a cluster of 14 genes, encoding the determinants of the flagellin glycosylation machinery in Pseudomonas aeruginosa PAK, which we called the flagellin glycosylation island. Flagellin glycosylation can be detected only in bacteria expressing the a-type flagellin sequence variants, and the survey of 30 P. aeruginosa isolates revealed coinheritance of the a-type flagellin genes with at least one of the flagellin glycosylation island genes. Expression of the b-type flagellin in PAK, an a-type strain carrying the glycosylation island, did not lead to glycosylation of the b-type flagellin of PAO1, suggesting that flagellins expressed by b-type bacteria not only lack the glycosylation island, they cannot serve as substrates for glycosylation. Providing the entire glycosylation island of PAK, including its a-type flagellin in a flagellin mutant of a b-type strain, results in glycosylation of the heterologous flagellin. These results suggest that some or all of the 14 genes on the glycosylation island are the genes that are missing from strain PAO1 to allow glycosylation of an appropriate flagellin. Inactivation of either one of the two flanking genes present on this island abolished flagellin glycosylation. Based on the limited homologies of these gene products with enzymes involved in glycosylation, we propose that the island encodes similar proteins involved in synthesis, activation, or polymerization of sugars that are necessary for flagellin glycosylation.
Figures
References
-
- Lechner J, Wieland F. Annu Rev Biochem. 1989;58:173–194. - PubMed
-
- Stimson E, Makepeace K, Dell A, Morris H R, Payne G, Saunders J R. Mol Microbiol. 1995;17:1201–1214. - PubMed
-
- Parge H E, Forest K T, Hickey M J, Christensen D A, Getzoff E D, Tainer J A. Nature (London) 1995;378:32–38. - PubMed
-
- Castric P. Microbiology. 1995;141:1247–1254. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
