

Polymerization of the SAM domain of TEL in leukemogenesis and transcriptional repression
- PMID: 11483520
- PMCID: PMC149168
- DOI: 10.1093/emboj/20.15.4173
Polymerization of the SAM domain of TEL in leukemogenesis and transcriptional repression
Abstract
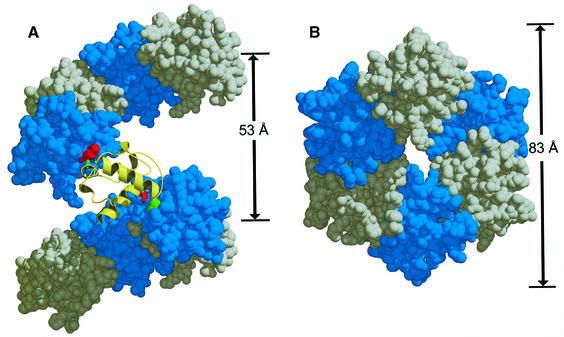
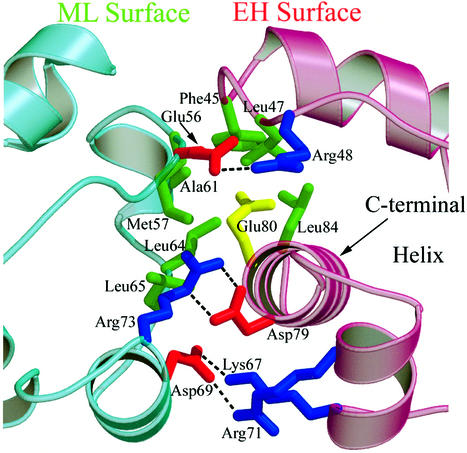
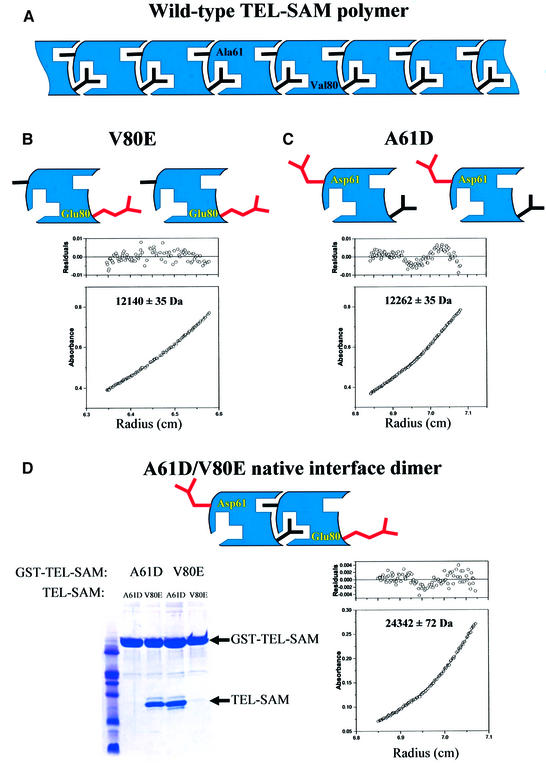
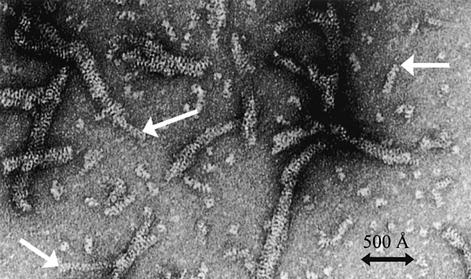
TEL is a transcriptional repressor that is a frequent target of chromosomal translocations in a large number of hematalogical malignancies. These rearrangements fuse a potent oligomerization module, the SAM domain of TEL, to a variety of tyrosine kinases or transcriptional regulatory proteins. The self-associating property of TEL-SAM is essential for cell transformation in many, if not all of these diseases. Here we show that the TEL-SAM domain forms a helical, head-to-tail polymeric structure held together by strong intermolecular contacts, providing the first clear demonstration that SAM domains can polymerize. Our results also suggest a mechanism by which SAM domains could mediate the spreading of transcriptional repression complexes along the chromosome.
Figures
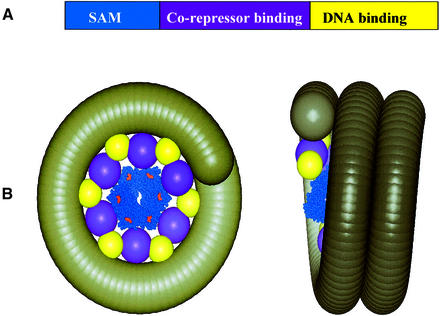
References
-
- Baker D.A., Mille-Baker,B., Wainwright,S.M., Ish-Horowicz,D. and Dibb,N.J. (2001) Mae mediates MAP kinase phosphorylation of Ets transcription factors in Drosophila. Nature, 411, 330–334. - PubMed
-
- Brunger A.T. et al. (1998) Crystallography & NMR system: a new software suite for macromolecular structure determination. Acta Crystallogr. D, 54, 905–921. - PubMed
-
- Carroll M., Tomasson,M.H., Barker,G.F., Golub,T.R. and Gilliland,D.G. (1996) The TEL/platelet-derived growth factor β receptor (PDGF β R) fusion in chronic myelomonocytic leukemia is a transforming protein that self-associates and activates PDGF β R kinase-dependent signaling pathways. Proc. Natl Acad. Sci. USA, 93, 14845–14850. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
