

Excision of deaminated cytosine from the vertebrate genome: role of the SMUG1 uracil-DNA glycosylase
- PMID: 11483530
- PMCID: PMC149160
- DOI: 10.1093/emboj/20.15.4278
Excision of deaminated cytosine from the vertebrate genome: role of the SMUG1 uracil-DNA glycosylase
Abstract
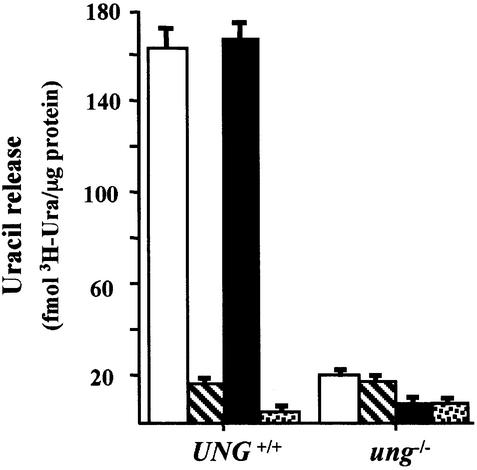
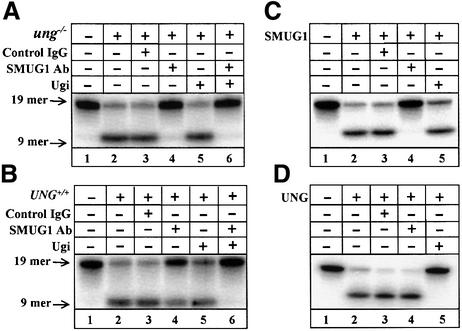
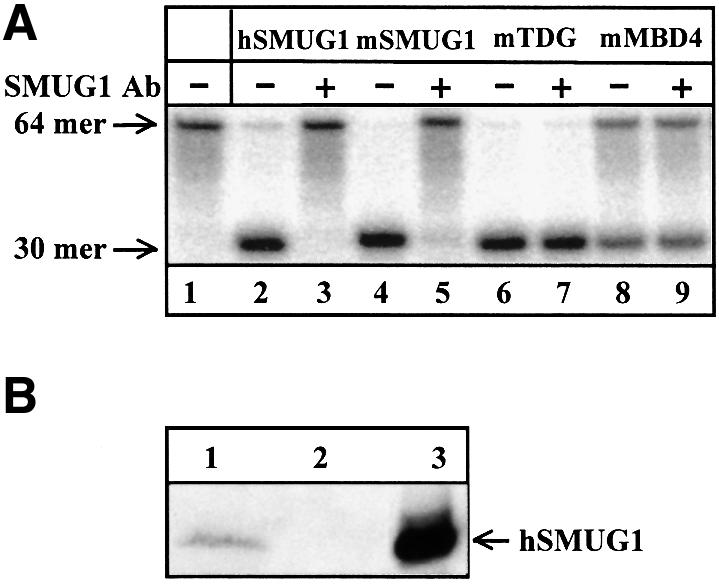
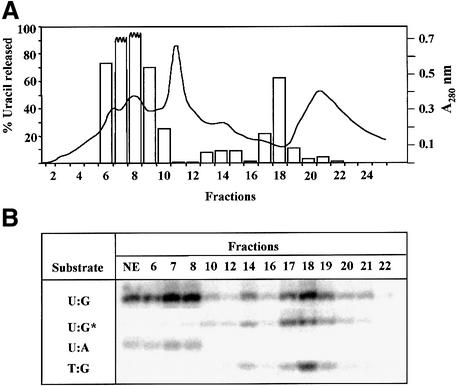
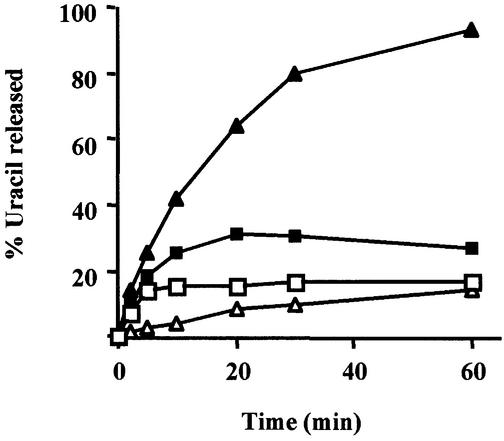
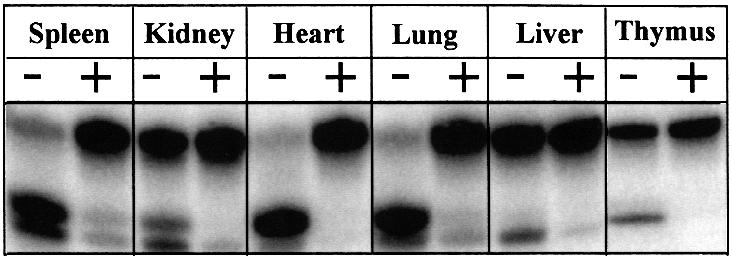
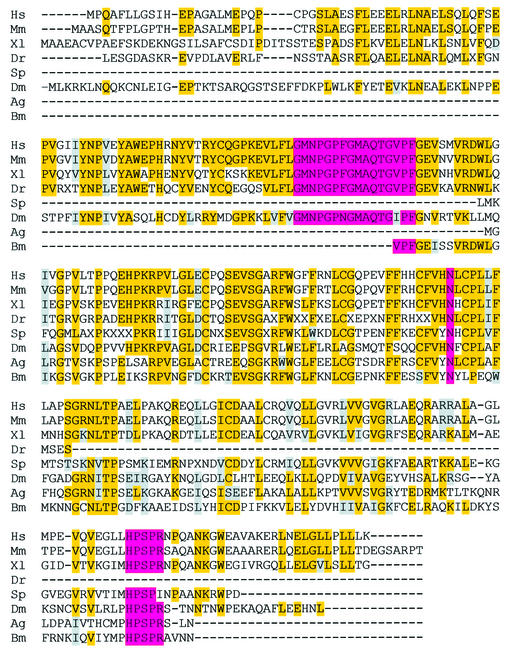
Gene-targeted mice deficient in the evolutionarily conserved uracil-DNA glycosylase encoded by the UNG gene surprisingly lack the mutator phenotype characteristic of bacterial and yeast ung(-) mutants. A complementary uracil-DNA glycosylase activity detected in ung(-/-) murine cells and tissues may be responsible for the repair of deaminated cytosine residues in vivo. Here, specific neutralizing antibodies were used to identify the SMUG1 enzyme as the major uracil-DNA glycosylase in UNG-deficient mice. SMUG1 is present at similar levels in cell nuclei of non-proliferating and proliferating tissues, indicating a replication- independent role in DNA repair. The SMUG1 enzyme is found in vertebrates and insects, whereas it is absent in nematodes, plants and fungi. We propose a model in which SMUG1 has evolved in higher eukaryotes as an anti-mutator distinct from the UNG enzyme, the latter being largely localized to replication foci in mammalian cells to counteract de novo dUMP incorporation into DNA.
Figures
References
-
- Aprelikova O.N. and Tomilin,N.V. (1982) Activity of uracil–DNA glycosylase in different rat tissues and in regenerating rat liver. FEBS Lett., 137, 193–195. - PubMed
-
- Brynolf K., Eliasson,R. and Reichard,P. (1978) Formation of Okazaki fragments in polyoma DNA synthesis caused by misincorporation of uracil. Cell, 13, 573–580. - PubMed
-
- Duncan B.K. and Miller,J.H. (1980) Mutagenic deamination of cytosine residues in DNA. Nature, 287, 560–561. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
