

Posttranslational processing of infected cell proteins 0 and 4 of herpes simplex virus 1 is sequential and reflects the subcellular compartment in which the proteins localize
- PMID: 11483735
- PMCID: PMC115034
- DOI: 10.1128/jvi.75.17.7904-7912.2001
Posttranslational processing of infected cell proteins 0 and 4 of herpes simplex virus 1 is sequential and reflects the subcellular compartment in which the proteins localize
Abstract
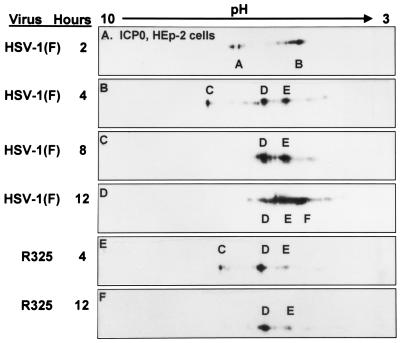
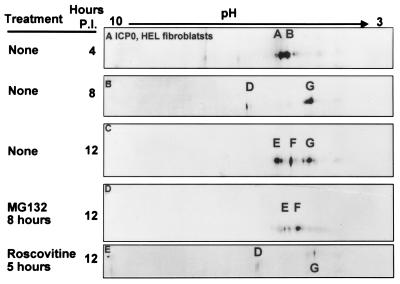
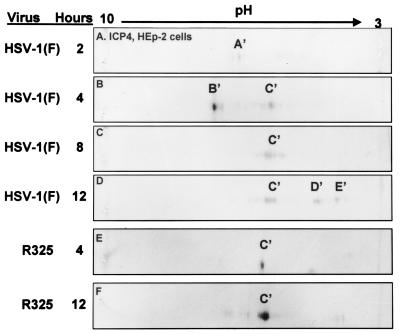
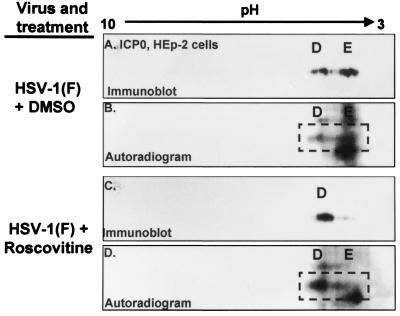
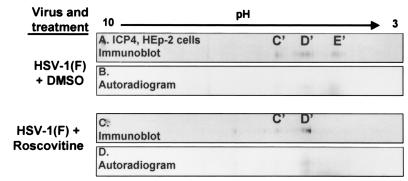
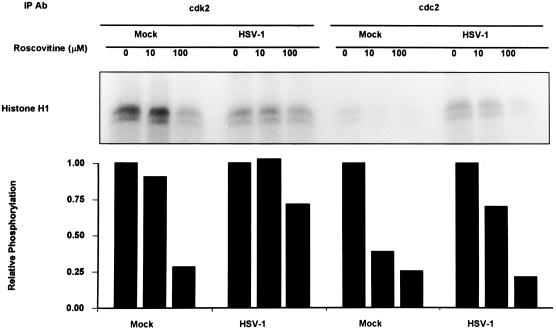
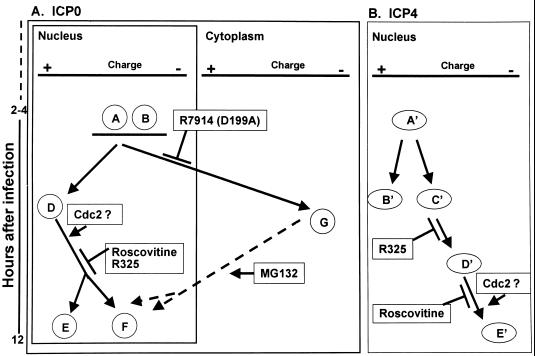
The herpes simplex virus 1 (HSV-1) infected cell proteins 0 and 4 (ICP0 and ICP4) are multifunctional proteins extensively posttranscriptionally processed by both cellular and viral enzymes. We examined by two-dimensional separations the posttranslational forms of ICP0 and ICP4 in HEp-2 cells and in human embryonic lung (HEL) fibroblasts infected with wild-type virus, mutant R325, lacking the sequences encoding the U(S)1.5 protein and the overlapping carboxyl-terminal domain of ICP22, or R7914, in which the aspartic acid 199 of ICP0 was replaced by alanine. We report the following (i) Both ICP0 and ICP4 were sequentially posttranslationally modified at least until 12 h after infection. In HEL fibroblasts, the processing of ICP0 shifted from A+B forms at 4 h to D+G forms at 8 h and finally to G, E, and F forms at 12 h. The ICP4 progression was from the A' form noted at 2 h to B' and C' forms noted at 4 h to the additional D' and E' forms noted at 12 h. The progression tended to be toward more highly charged forms of the proteins. (ii) Although the overall patterns were similar, the mobility of proteins made in HEp-2 cells differed from those made in HEL fibroblasts. (iii) The processing of ICP0 forms E and F was blocked in HEL fibroblasts infected with R325 or with wild-type virus and treated with roscovitine, a specific inhibitor of cell cycle-dependent kinases cdc2, cdk2, and cdk5. R325-infected HEp-2 cells lacked the D' form of ICP4, and roscovitine blocked the appearance of the most highly charged E' form of ICP4. (iv) A characteristic of ICP0 is that it is translocated into the cytoplasm of HEL fibroblasts between 5 and 9 h after infection. Addition of MG132 to the cultures late in infection resulted in rapid relocation of cytoplasmic ICP0 back into the nucleus. Exposure of HEL fibroblasts to MG132 late in infection resulted in the disappearance of the highly charged ICP0 G isoform. The G form of ICP0 was also absent in cells infected with R7914 mutant. In cells infected with this mutant, ICP0 is not translocated to the cytoplasm. (v) Last, cdc2 was active in infected cells, and this activity was inhibited by roscovitine. In contrast, the activity of cdk2 exhibited by immunoprecipitated protein was reduced and resistant to roscovitine and may represent a contaminating kinase activity. We conclude from these results that the ICP0 G isoform is the cytoplasmic form, that it may be phosphorylated by cdc2, consistent with evidence published earlier (S. J., Advani, R. R. Weichselbaum, and B. Roizman, Proc. Natl. Acad. Sci. USA 96:10996-11001, 2000), and that the processing is reversed upon relocation of the G isoform from the cytoplasm into the nucleus. The processing of ICP4 is also affected by R325 and roscovitine. The latter result suggests that ICP4 may also be a substrate of cdc2 late in infection. Last, additional modifications are superimposed by cell-type-specific enzymes.
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
