

Specific interaction of a novel foamy virus Env leader protein with the N-terminal Gag domain
- PMID: 11483744
- PMCID: PMC115043
- DOI: 10.1128/jvi.75.17.7995-8007.2001
Specific interaction of a novel foamy virus Env leader protein with the N-terminal Gag domain
Abstract
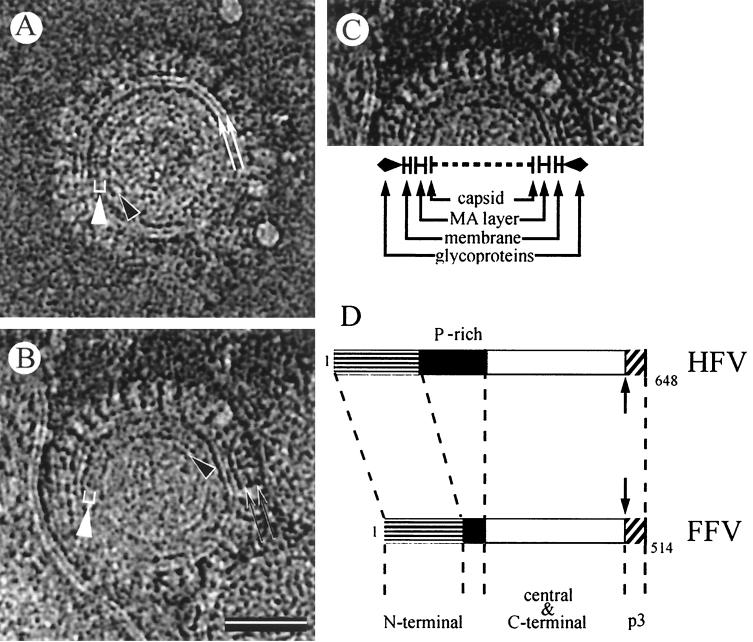
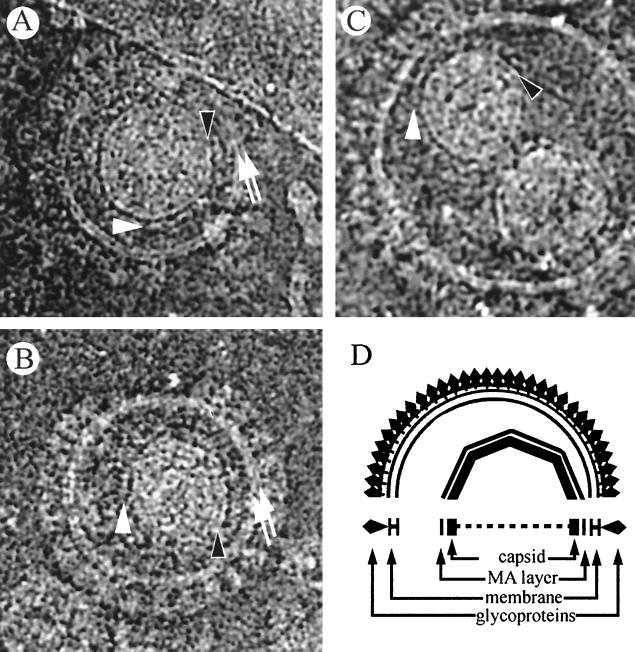
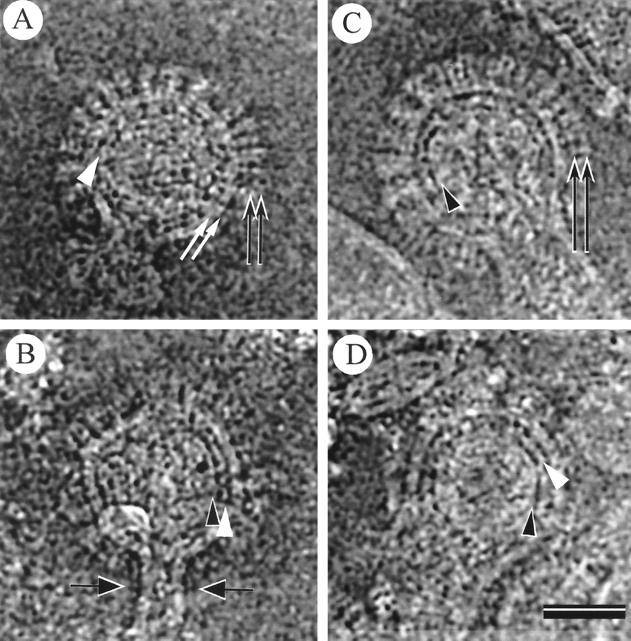
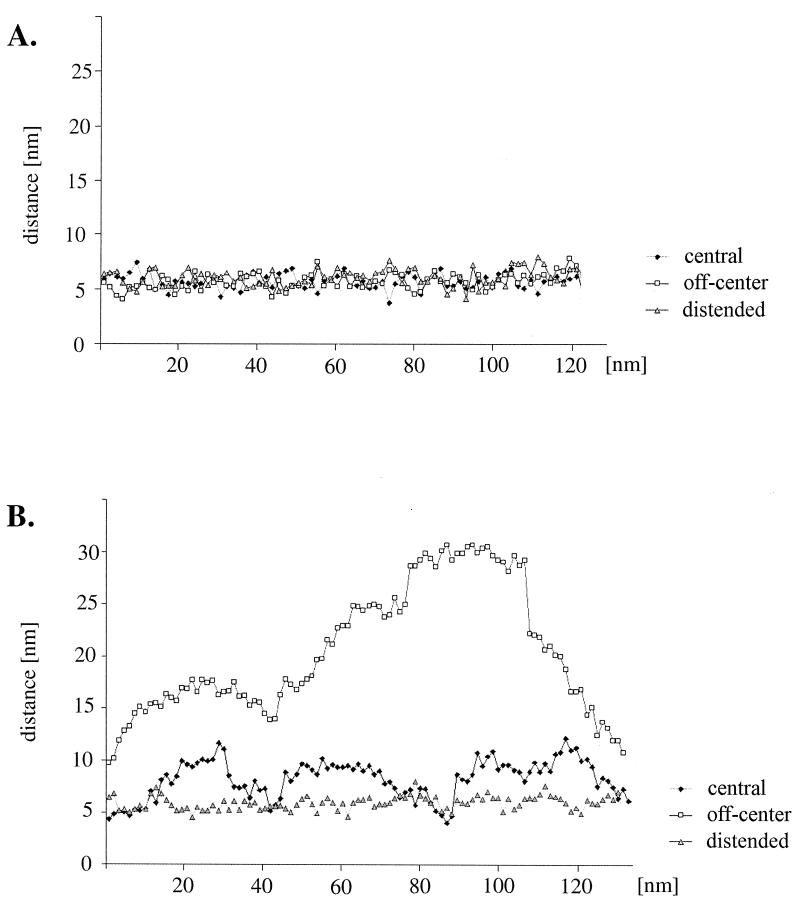
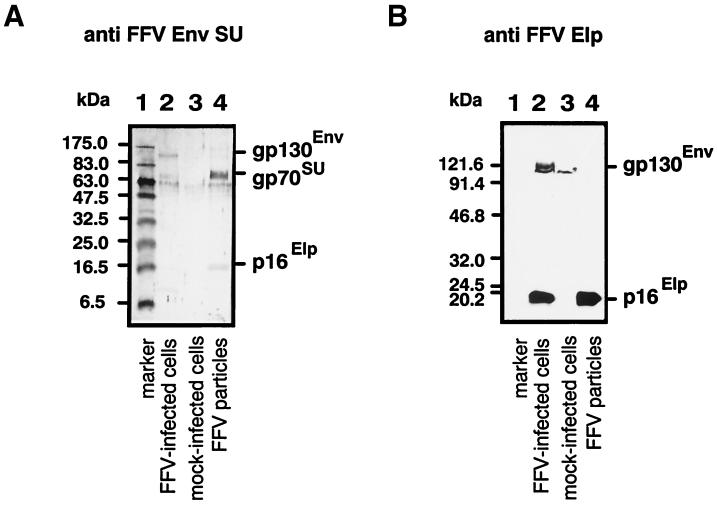
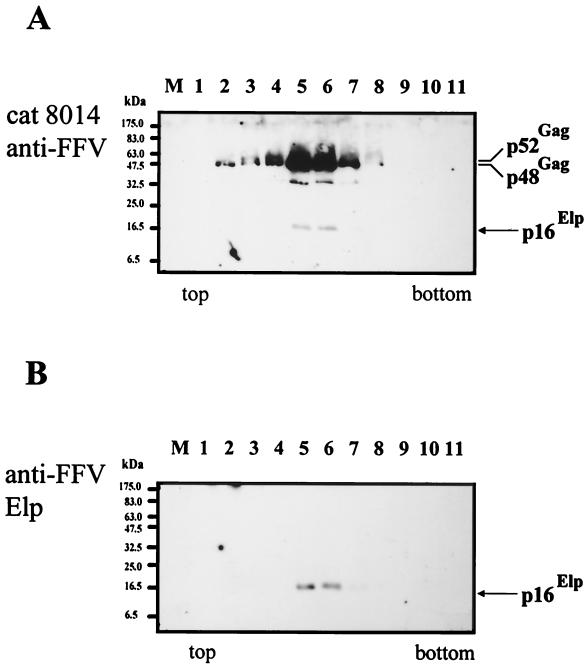
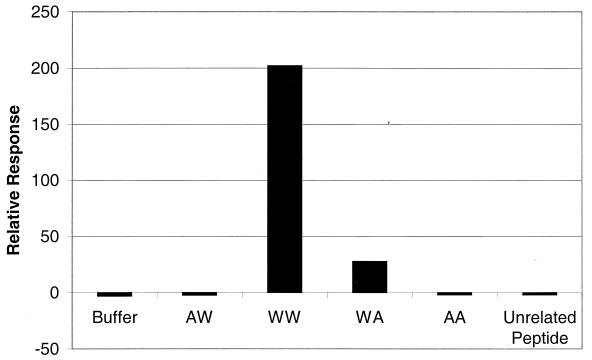
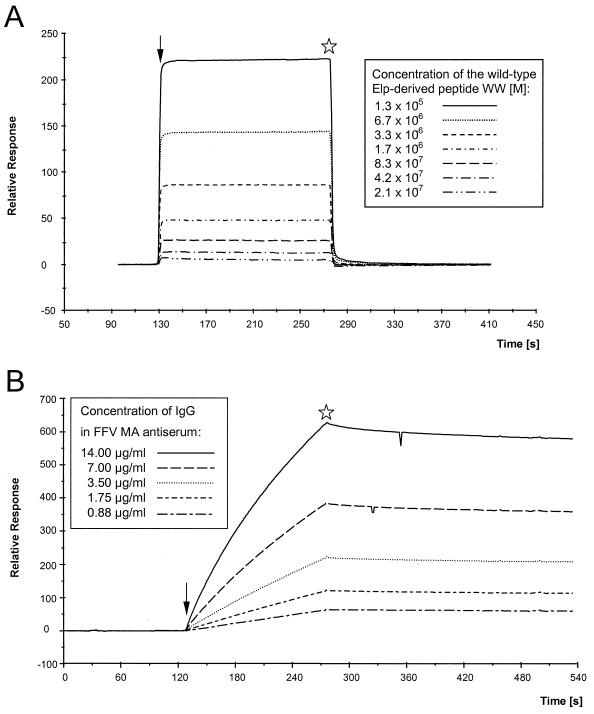
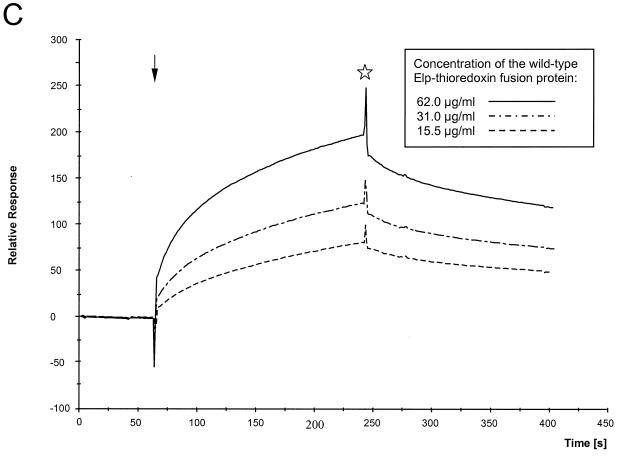
Cryoelectron micrographs of purified human foamy virus (HFV) and feline foamy virus (FFV) particles revealed distinct radial arrangements of Gag proteins. The capsids were surrounded by an internal Gag layer that in turn was surrounded by, and separated from, the viral membrane. The width of this layer was about 8 nm for HFV and 3.8 nm for FFV. This difference in width is assumed to reflect the different sizes of the HFV and FFV MA domains: the HFV MA domain is about 130 residues longer than that of FFV. The distances between the MA layer and the edge of the capsid were identical in different particle classes. In contrast, only particles with a distended envelope displayed an invariant, close spacing between the MA layer and the Env membrane which was absent in the majority of particles. This indicates a specific interaction between MA and Env at an unknown step of morphogenesis. This observation was supported by surface plasmon resonance studies. The purified N-terminal domain of FFV Gag specifically interacted with synthetic peptides and a defined protein domain derived from the N-terminal Env leader protein. The specificity of this interaction was demonstrated by using peptides varying in the conserved Trp residues that are known to be required for HFV budding. The interaction with Gag required residues within the novel virion-associated FFV Env leader protein of about 16.5 kDa.
Figures
References
-
- Alke A, Schwantes A, Zemba M, Flügel R M, Löchelt M. Characterization of the humoral immune response and virus replication in cats experimentally infected with feline foamy virus. Virology. 2000;275:170–176. - PubMed
-
- Bodem J, Zemba M, Flügel R M. Nuclear localization of the functional Bel 1 transactivator but not of the gag proteins of the feline foamy virus. Virology. 1998;251:22–27. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
