

Expression of the three human major histocompatibility complex class II isotypes exhibits a differential dependence on the transcription factor RFXAP
- PMID: 11486010
- PMCID: PMC87290
- DOI: 10.1128/MCB.21.17.5699-5709.2001
Expression of the three human major histocompatibility complex class II isotypes exhibits a differential dependence on the transcription factor RFXAP
Abstract
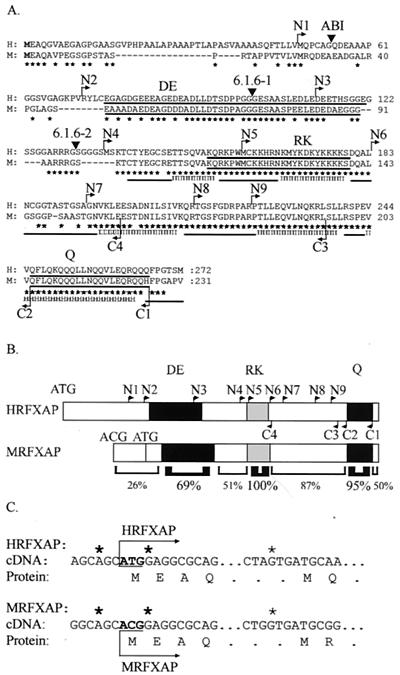
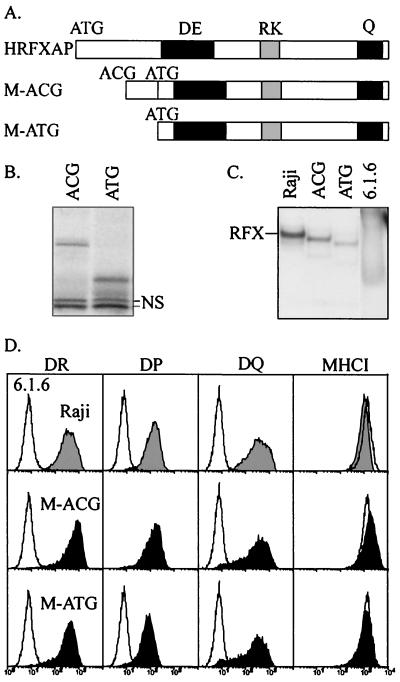
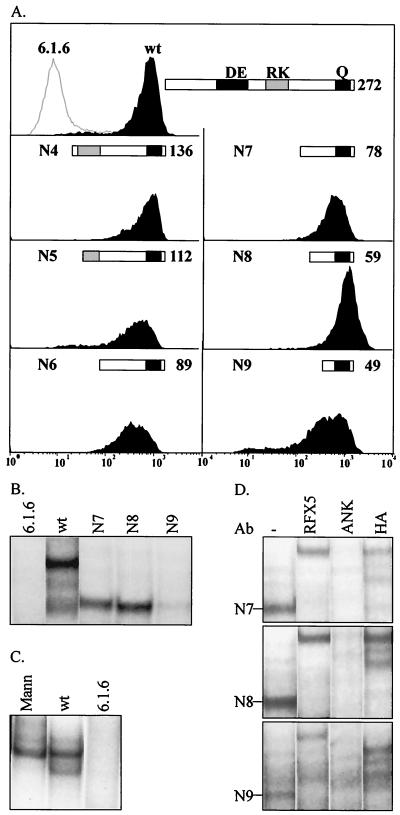
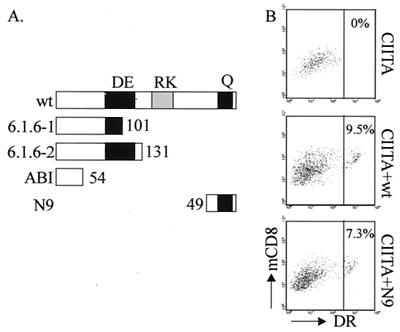
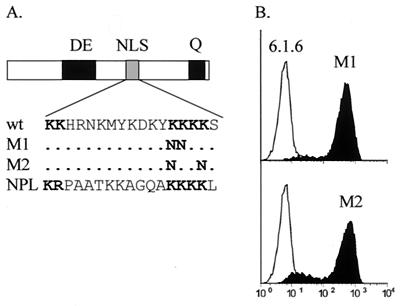
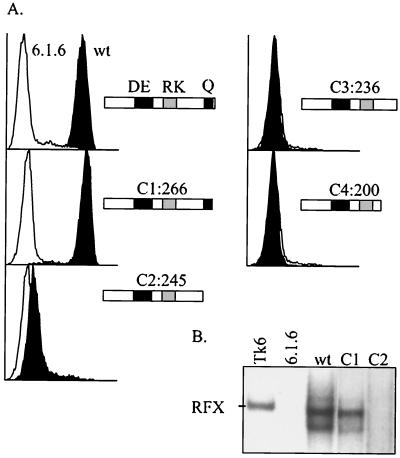
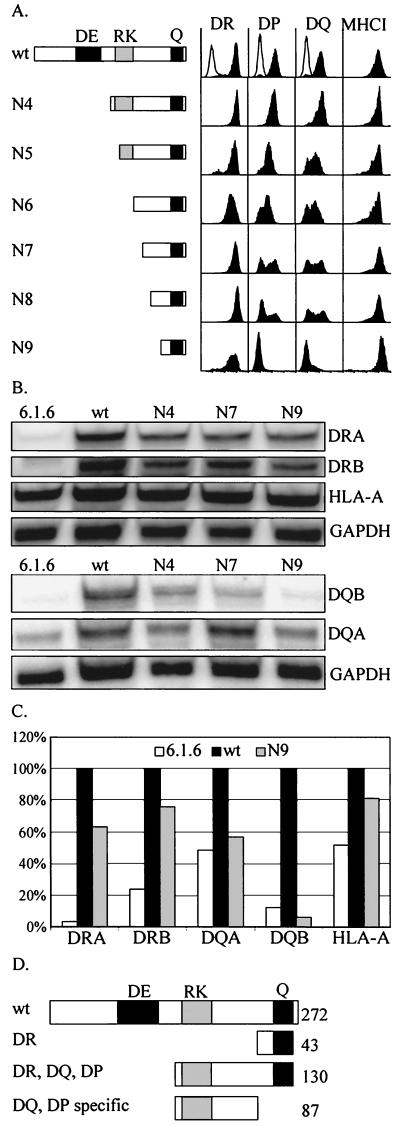
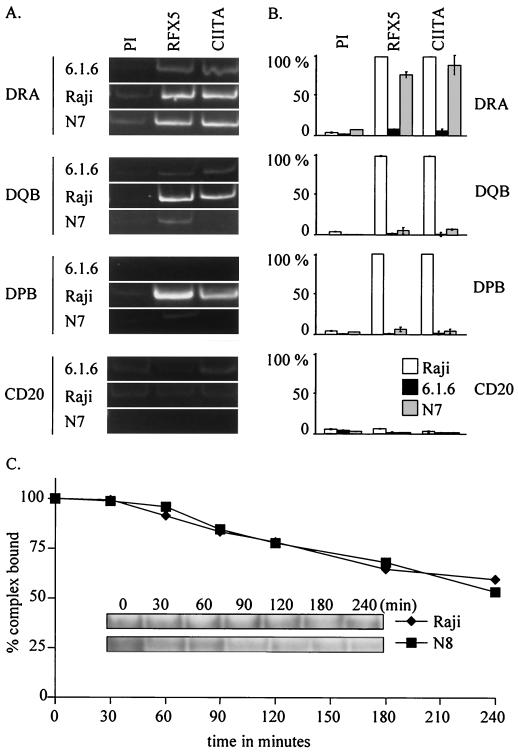
Major histocompatibility complex class II (MHCII) molecules play a pivotal role in the immune system because they direct the development and activation of CD4(+) T cells. There are three human MHCII isotypes, HLA-DR, HLA-DQ, and HLA-DP. Key transcription factors controlling MHCII genes have been identified by virtue of the fact that they are mutated in a hereditary immunodeficiency resulting from a lack of MHCII expression. RFXAP-one of the factors affected in this disease-is a subunit of RFX, a DNA-binding complex that recognizes the X box present in all MHCII promoters. To facilitate identification of conserved regions in RFXAP, we isolated the mouse gene. We then delimited conserved domains required to restore endogenous MHCII expression in cell lines lacking a functional RFXAP gene. Surprisingly, we found that 80% of RFXAP is dispensable for the reactivation of DR expression. Only a short C-terminal segment of the protein is essential for this isotype. In contrast, optimal expression of DQ and DP requires a larger C-terminal segment. These results define an RFXAP domain with an MHCII isotype-specific function. Expression of the three MHCII isotypes exhibits a differential requirement for this domain. We show that this is due to a differential dependence on this domain for promoter occupation and recruitment of the coactivator CIITA in vivo.
Figures
References
-
- Alcaide-Loridan C, Lennon A M, Bono M R, Barbouche R, Dellagi K, Fellous M. Differential expression of MHC class II isotype chains. Microbes Infect. 1999;1:929–934. - PubMed
-
- Boss J M. Regulation of transcription of MHC class II genes. Curr Opin Immunol. 1997;9:107–113. - PubMed
-
- Caretti G, Cocchiarella F, Sidoli C, Villard J, Peretti M, Reith W, Mantovani R. Dissection of functional NF-Y–RFX cooperative interactions on the MHC class II E alpha promoter. J Biol Chem. 2000;302:539–552. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials