

Induction of cyclin D1 transcription and CDK2 activity by Notch(ic): implication for cell cycle disruption in transformation by Notch(ic)
- PMID: 11486031
- PMCID: PMC87311
- DOI: 10.1128/MCB.21.17.5925-5934.2001
Induction of cyclin D1 transcription and CDK2 activity by Notch(ic): implication for cell cycle disruption in transformation by Notch(ic)
Abstract
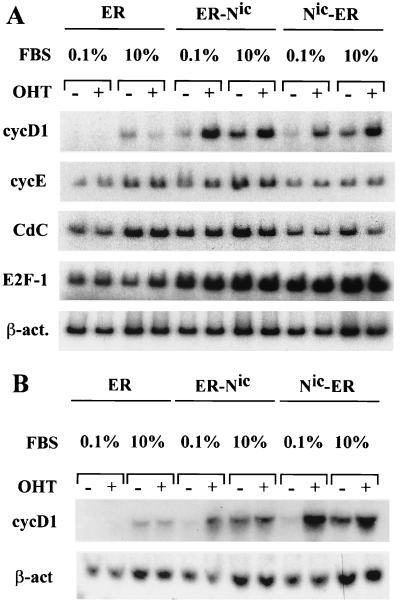
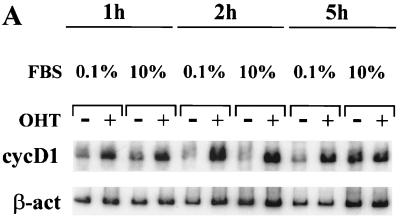
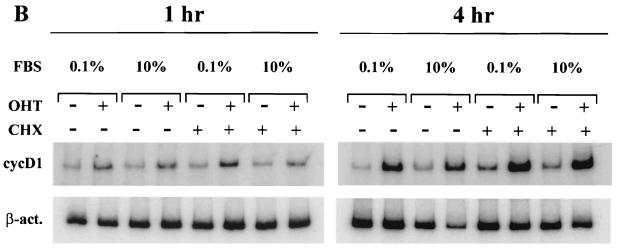
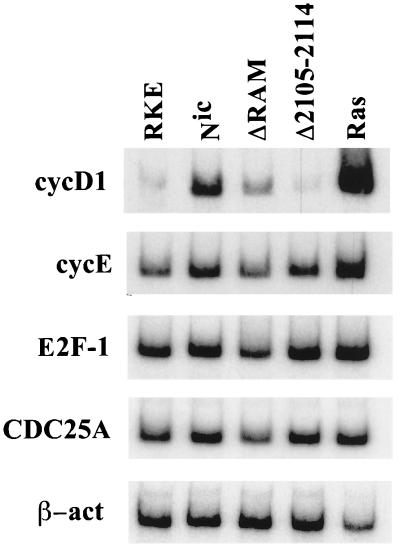
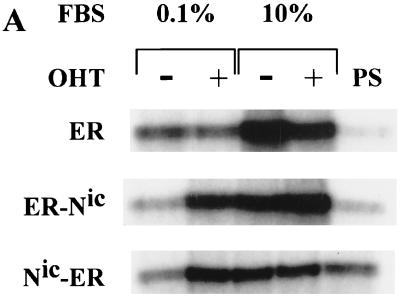
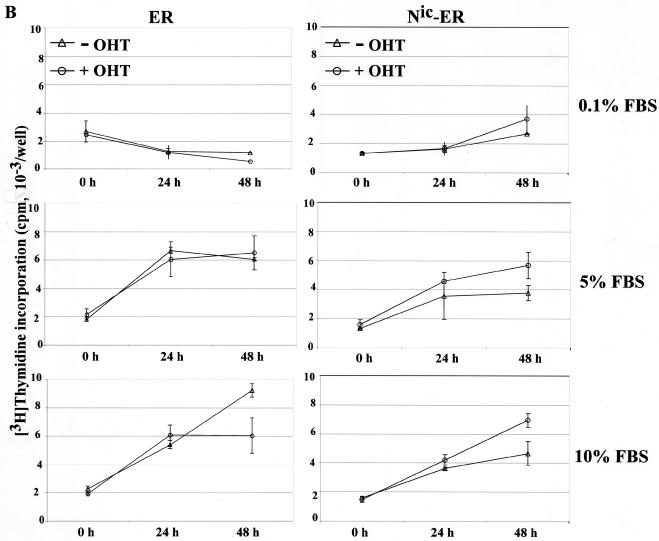
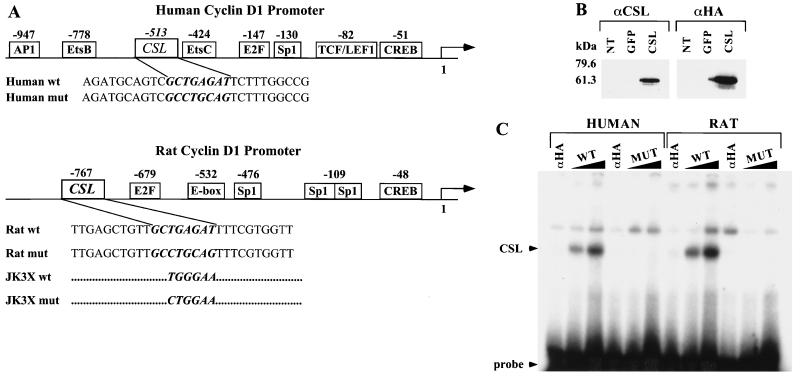
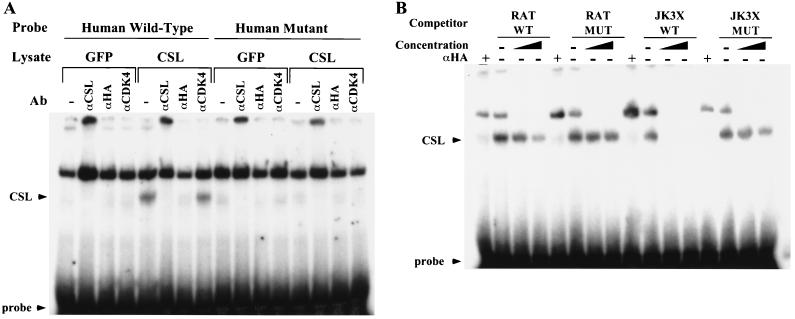
Notch genes encode a family of transmembrane proteins that are involved in many cellular processes such as differentiation, proliferation, and apoptosis. Although it is well established that all four Notch genes can act as oncogenes, the mechanism by which Notch proteins transform cells remains unknown. Previously, we have shown that transformation of RKE cells can be conditionally induced by hormone activation of Notch(ic)-estrogen receptor (ER) chimeras. Using this inducible system, we show that Notch(ic) activates transcription of the cyclin D1 gene with rapid kinetics. Transcriptional activation of cyclin D1 is independent from serum-derived growth factors and de novo synthesis of secondary transcriptional activators. Moreover, hormone activation of Notch(ic)-ER proteins induces CDK2 activity in the absence of serum. Upregulation of cyclin D1 and activation of CDK2 by Notch(ic) result in the promotion of S-phase entry. These data demonstrate the first evidence that Notch(ic) proteins can directly regulate factors involved in cell cycle control and affect cellular proliferation. Furthermore, nontransforming Notch(ic) proteins do not induce cyclin D1 expression, indicating that the mechanism of transformation involves cell cycle deregulation through constitutive expression of cyclin D1. Finally, we have identified a CSL [stands for CBF1, Su(H), and Lag-1] binding site within the human and rat cyclin D1 promoters, suggesting that Notch(ic) proteins activate cyclin D1 transcription through a CSL-dependent pathway.
Figures
Similar articles
-
Characterization of a high-molecular-weight Notch complex in the nucleus of Notch(ic)-transformed RKE cells and in a human T-cell leukemia cell line.Mol Cell Biol. 2002 Jun;22(11):3927-41. doi: 10.1128/MCB.22.11.3927-3941.2002. Mol Cell Biol. 2002. PMID: 11997524 Free PMC article.
-
c-Myc or cyclin D1 mimics estrogen effects on cyclin E-Cdk2 activation and cell cycle reentry.Mol Cell Biol. 1998 Aug;18(8):4499-508. doi: 10.1128/MCB.18.8.4499. Mol Cell Biol. 1998. PMID: 9671459 Free PMC article.
-
Notch(ic)-ER chimeras display hormone-dependent transformation, nuclear accumulation, phosphorylation and CBF1 activation.Oncogene. 2000 Aug 10;19(34):3914-24. doi: 10.1038/sj.onc.1203719. Oncogene. 2000. PMID: 10951584
-
Multifaceted regulation of cell cycle progression by estrogen: regulation of Cdk inhibitors and Cdc25A independent of cyclin D1-Cdk4 function.Mol Cell Biol. 2001 Feb;21(3):794-810. doi: 10.1128/MCB.21.3.794-810.2001. Mol Cell Biol. 2001. PMID: 11154267 Free PMC article.
-
Estrogen and antiestrogen regulation of cell cycle progression in breast cancer cells.Endocr Relat Cancer. 2003 Jun;10(2):179-86. doi: 10.1677/erc.0.0100179. Endocr Relat Cancer. 2003. PMID: 12790780 Review.
Cited by
-
ASPM stabilizes the NOTCH intracellular domain 1 and promotes oncogenesis by blocking FBXW7 binding in hepatocellular carcinoma cells.Mol Oncol. 2024 Mar;18(3):562-579. doi: 10.1002/1878-0261.13589. Epub 2024 Jan 26. Mol Oncol. 2024. PMID: 38279565 Free PMC article.
-
Notch activation induces endothelial cell cycle arrest and participates in contact inhibition: role of p21Cip1 repression.Mol Cell Biol. 2004 Oct;24(20):8813-22. doi: 10.1128/MCB.24.20.8813-8822.2004. Mol Cell Biol. 2004. PMID: 15456857 Free PMC article.
-
Notch pathway molecules are essential for the maintenance, but not the generation, of mammalian neural stem cells.Genes Dev. 2002 Apr 1;16(7):846-58. doi: 10.1101/gad.975202. Genes Dev. 2002. PMID: 11937492 Free PMC article.
-
NOTCH1 signaling contributes to cell growth, anti-apoptosis and metastasis in salivary adenoid cystic carcinoma.Oncotarget. 2014 Aug 30;5(16):6885-95. doi: 10.18632/oncotarget.2321. Oncotarget. 2014. PMID: 25149541 Free PMC article.
-
Cancer stem cells: cell culture, markers, and targets for new therapies.J Cell Biochem. 2009 Dec 1;108(5):1031-8. doi: 10.1002/jcb.22350. J Cell Biochem. 2009. PMID: 19760641 Free PMC article. Review.
References
-
- Albanese C, Johnson J, Watanabe G, Eklund N, Vu D, Arnold A, Pestell R G. Transforming p21ras mutants and c-Ets-2 activate the cyclin D1 promoter through distinguishable regions. J Biol Chem. 1995;270:23589–23597. - PubMed
-
- Aster J, Pear W, Hasserjian R, Erba H, Davi F, Luo B, Scott M, Baltimore D, Sklar J. Functional analysis of the TAN-1 gene, a human homolog of Drosophila notch. Cold Spring Harbor Symp Quant Biol. 1994;59:125–136. - PubMed
-
- Bellavia D, Campese A F, Alesse E, Vacca A, Felli M P, Balestri A, Stoppacciaro A, Tiveron C, Tatangelo L, Giovarelli M, Gaetano C, Ruco L, Hoffman E S, Hayday A C, Lendahl U, Frati L, Gulino A, Screpanti I. Constitutive activation of NF-kappaB and T-cell leukemia/lymphoma in Notch3 transgenic mice. EMBO J. 2000;19:3337–3348. - PMC - PubMed
-
- Bromberg J F, Wrzeszczynska M H, Devgan G, Zhao Y, Pestell R G, Albanese C, Darnell J E., Jr Stat3 as an oncogene. Cell. 1999;98:295–303. . (Erratum, 99:239.) - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials