

Altered processing of pro-orphanin FQ/nociceptin and pro-opiomelanocortin-derived peptides in the brains of mice expressing defective prohormone convertase 2
- PMID: 11487609
- PMCID: PMC6763138
- DOI: 10.1523/JNEUROSCI.21-16-05864.2001
Altered processing of pro-orphanin FQ/nociceptin and pro-opiomelanocortin-derived peptides in the brains of mice expressing defective prohormone convertase 2
Abstract
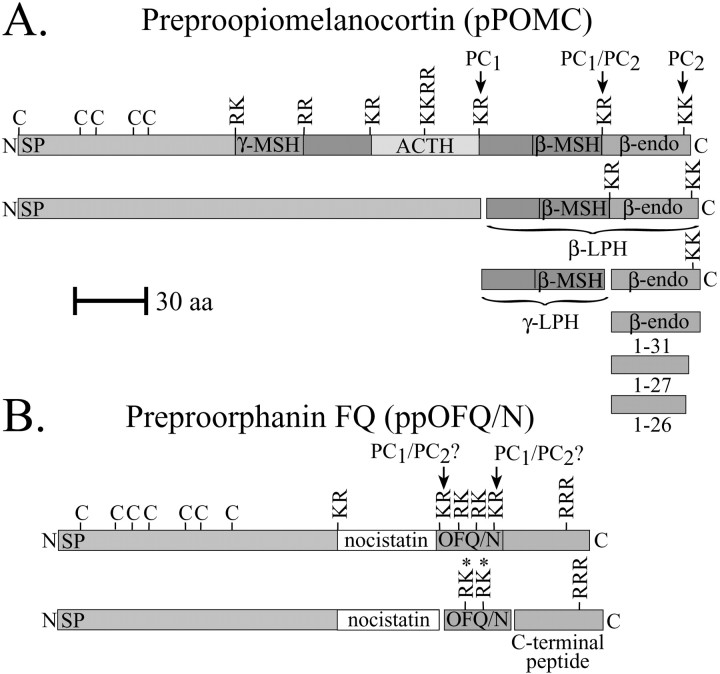
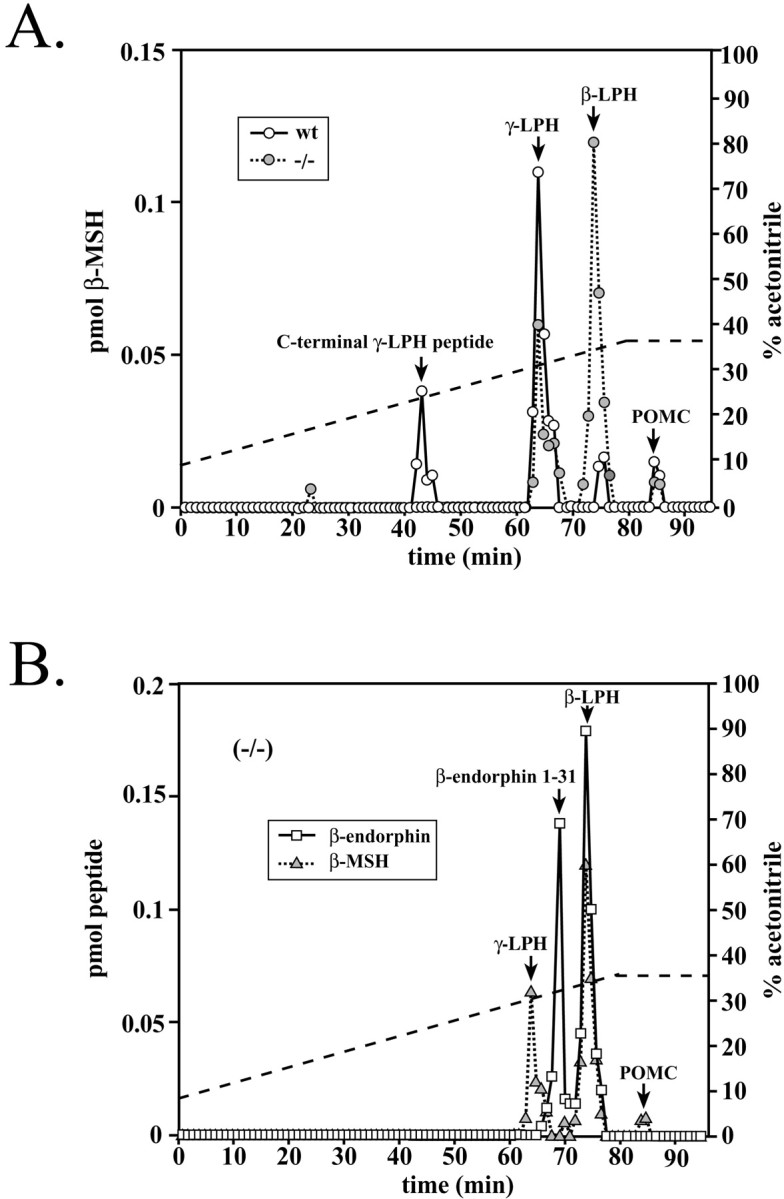
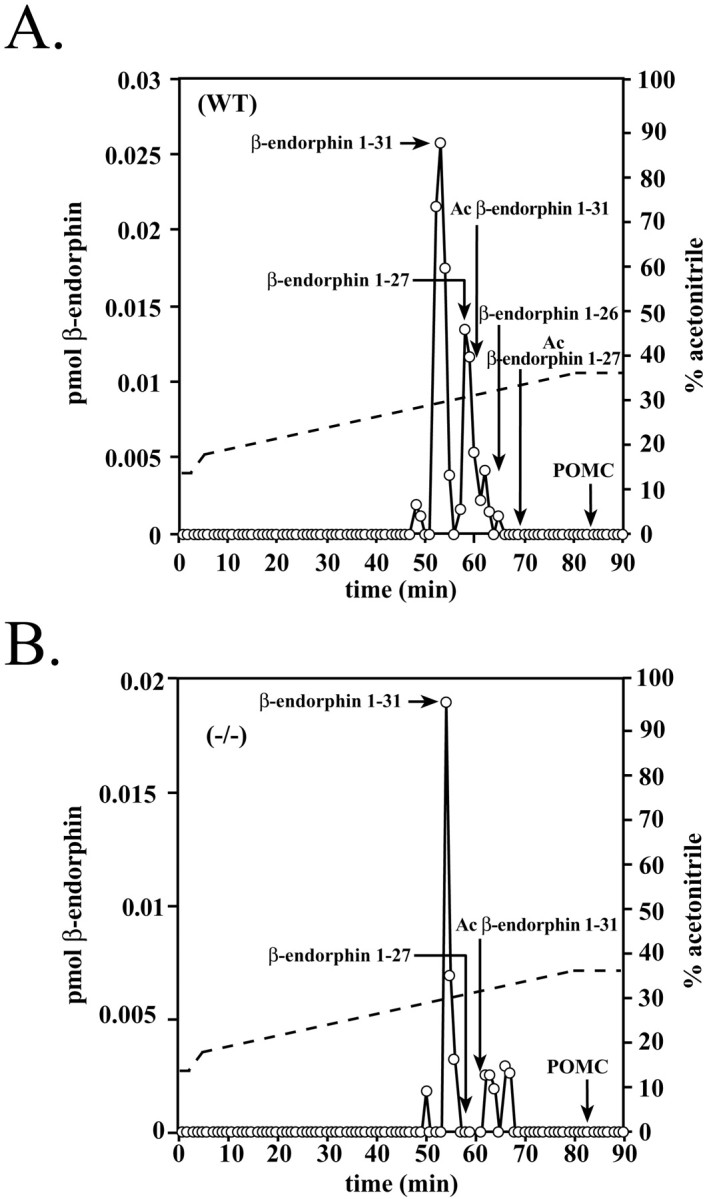
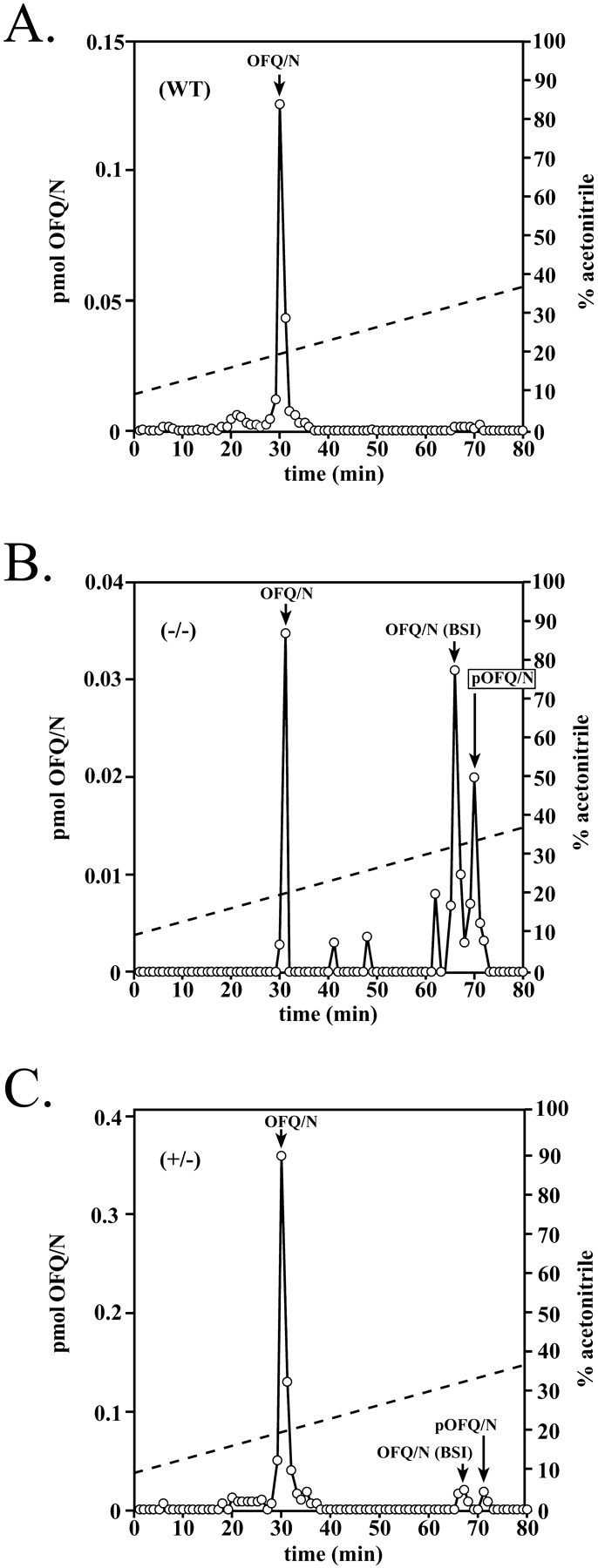
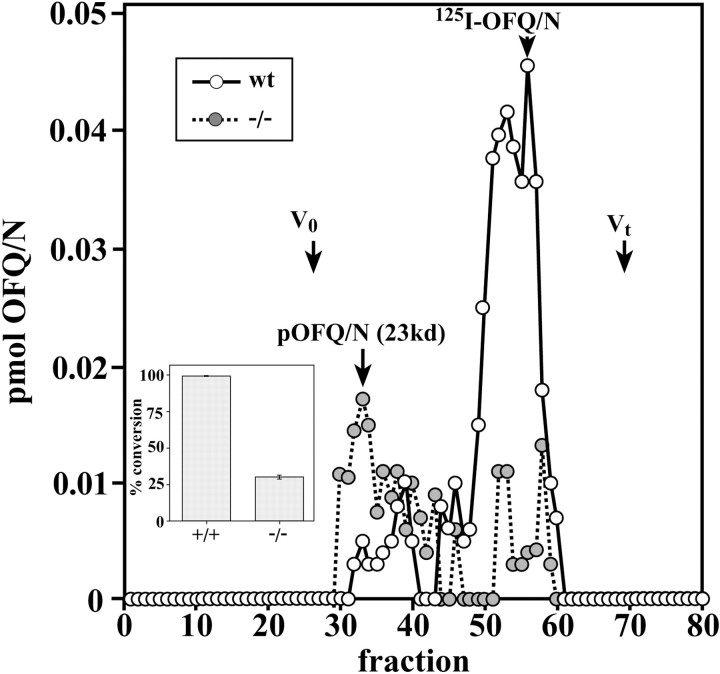
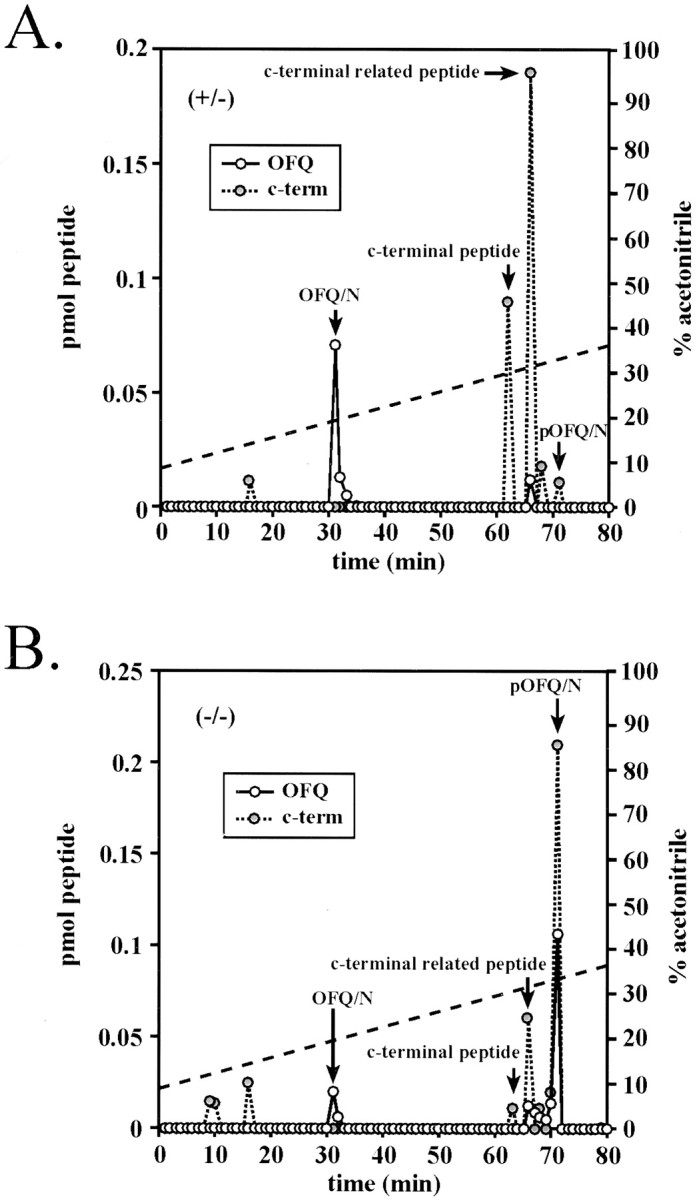
The bioactivity of neuropeptides can be regulated by a variety of post-translational modifications, including proteolytic processing. Here, gene-targeted mice producing defective prohormone convertase 2 (PC2) were used to examine the post-translational processing of two neuroendocrine prohormones, pro-opiomelanocortin (POMC) and pro-orphanin FQ (pOFQ)/nociceptin (N), in the brain. Reversed-phase HPLC and gel-exclusion chromatography were combined with specific radioimmunoassays to analyze the processing patterns of these two prohormones in the hypothalamus and the amygdala. In the case of POMC, the lack of PC2 activity completely prevented carboxy-shortening of beta-endorphins and greatly diminished conversion of beta-lipotropin to gamma-lipotropin and beta-endorphin. Although conversion of beta-lipotropin to beta-endorphin decreased, the lack of PC2 activity caused an increase in beta-lipotropin and beta-endorphin levels in the mutant animals, but no increases in POMC or biosynthetic intermediates were seen. The extent of OFQ/N production was significantly lower in PC2-deficient mice and there was an accumulation of relatively large amounts of pOFQ/N and biosynthetic intermediates. These results demonstrate that PC2 is directly involved in the biogenesis of two brain neuropeptides in vivo and suggest that the specific prohormone and cellular context influences neuropeptide processing by PCs.
Figures


References
-
- Allen RG, Pellegrino MJ, Peng, B, Pintar JE (1999) Altered POMC processing in SPC2 KO mice. Program Abstr Endocr Soc Annu Meet 270.99.
-
- Berman Y, Mzhavia N, Polonskaia A, Furuta M, Steiner D, Pintar J, Devi L. Defective prodynorphin processing in mice lacking prohormone convertase PC2. J Neurochem. 2000;75:1763–1770. - PubMed
-
- Bunzow JR, Saez C, Mortrud M, Bouvier C, Williams JT, Low M, Grandy DK. Molecular cloning and tissue distribution of a putative member of the rat opioid receptor gene family that is not a μ, δ, or κ opioid receptor type. FEBS Lett. 1994;347:284–288. - PubMed
-
- Day R, Schafer M, Watson SJ, Chretien M, Seidah NG. Distribution and regulation of the prohormone convertases PC1 and PC2 in the rat pituitary. Mol Endocrinol. 1992;6:485–497. - PubMed
-
- Eipper BA, Mains RE. Structure and biosynthesis of pro-adrenocorticotropin/endorphin and related peptides. Endocr Rev. 1980;1:1–27. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous