

Molecular bases of odor discrimination: Reconstitution of olfactory receptors that recognize overlapping sets of odorants
- PMID: 11487625
- PMCID: PMC6763140
- DOI: 10.1523/JNEUROSCI.21-16-06018.2001
Molecular bases of odor discrimination: Reconstitution of olfactory receptors that recognize overlapping sets of odorants
Abstract
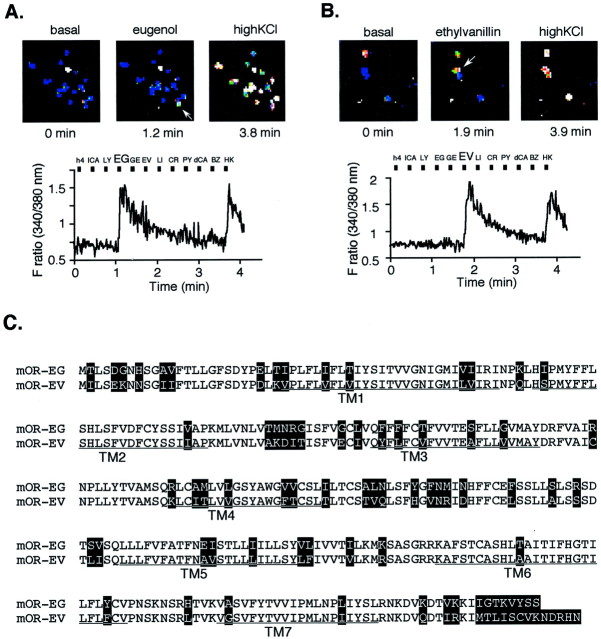
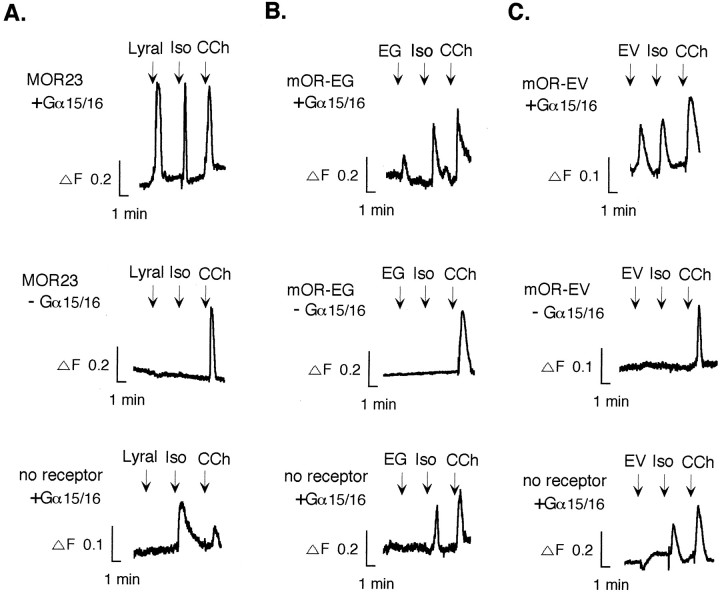
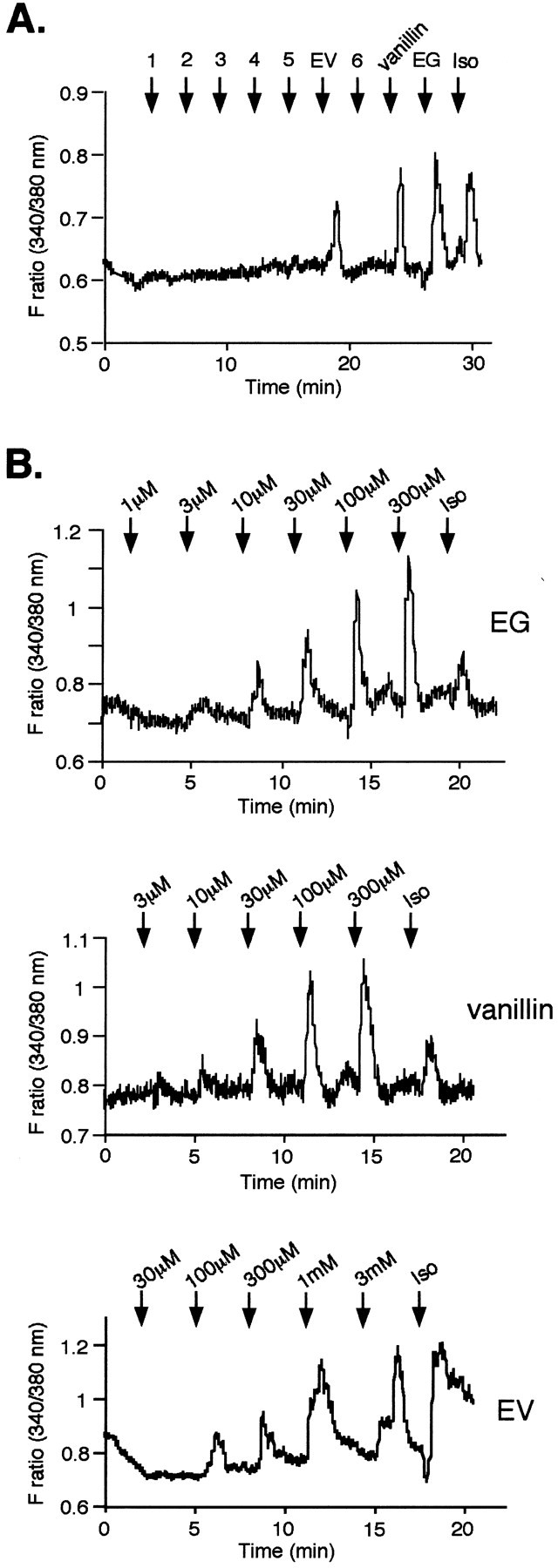
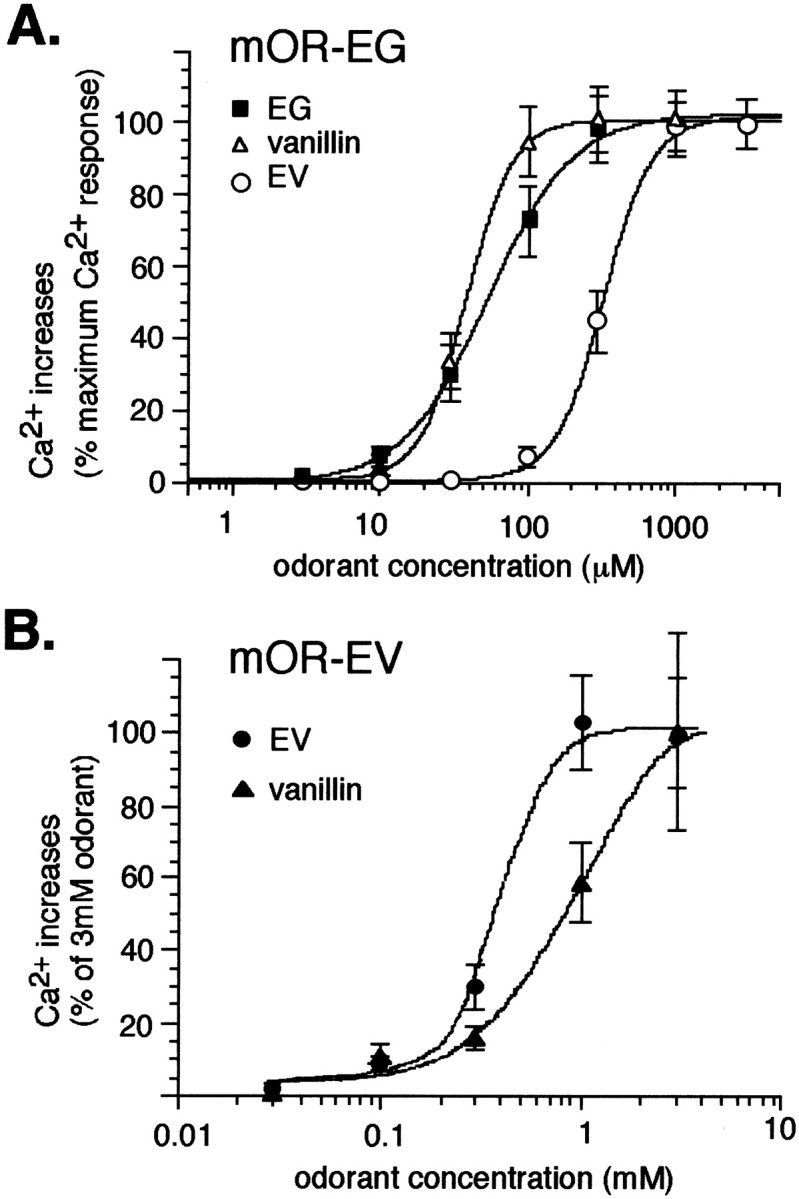
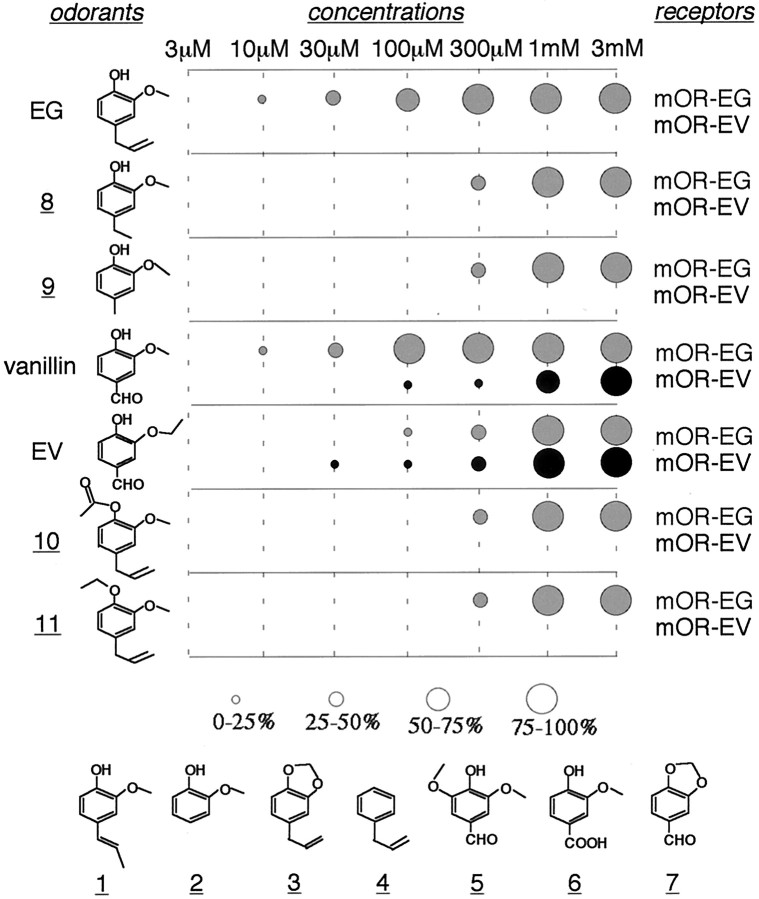
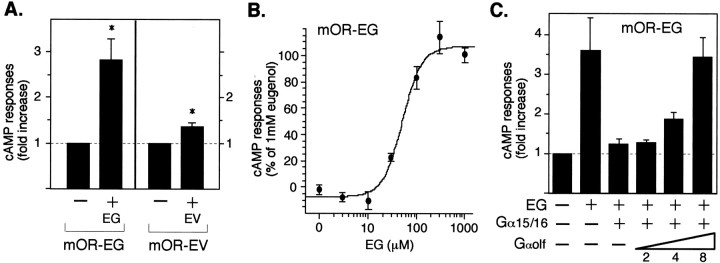
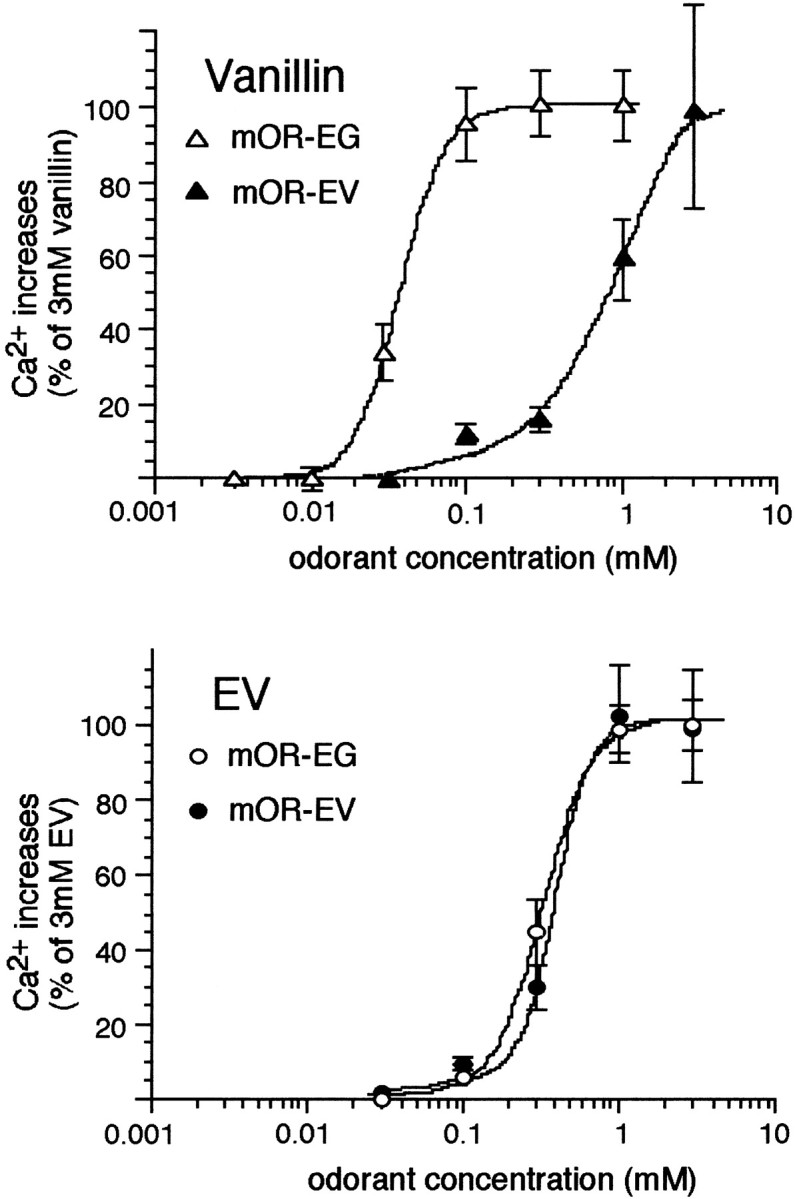
The vertebrate olfactory system discriminates a wide variety of odorants by relaying coded information from olfactory sensory neurons in the olfactory epithelium to olfactory cortical areas of the brain. Recent studies have shown that the first step in odor discrimination is mediated by approximately 1000 distinct olfactory receptors, which comprise the largest family of G-protein-coupled receptors. In the present study, we used Ca(2+) imaging and single-cell reverse transcription-PCR techniques to identify mouse olfactory neurons responding to an odorant and subsequently to clone a receptor gene from the responsive cell. The functionally cloned receptors were expressed in heterologous systems, demonstrating that structurally related olfactory receptors recognized overlapping sets of odorants with distinct affinities and specificities. Our results provide direct evidence for the existence of a receptor code in which the identities of different odorants are specified by distinct combinations of odorant receptors that possess unique molecular receptive ranges. We further demonstrate that the receptor code for an odorant changes with odorant concentration. Finally, we show that odorant receptors in human embryonic kidney 293 cells couple to stimulatory G-proteins such as Galphaolf, resulting in odorant-dependent increases in cAMP. Odor discrimination is thus determined by differences in the receptive ranges of the odorant receptors that together encode specific odorant molecules.
Figures
Similar articles
-
Odor discrimination by G protein-coupled olfactory receptors.Microsc Res Tech. 2002 Aug 1;58(3):135-41. doi: 10.1002/jemt.10131. Microsc Res Tech. 2002. PMID: 12203691 Review.
-
Olfactory receptor antagonism between odorants.EMBO J. 2004 Jan 14;23(1):120-6. doi: 10.1038/sj.emboj.7600032. Epub 2003 Dec 18. EMBO J. 2004. PMID: 14685265 Free PMC article.
-
Functional identification and reconstitution of an odorant receptor in single olfactory neurons.Proc Natl Acad Sci U S A. 1999 Mar 30;96(7):4040-5. doi: 10.1073/pnas.96.7.4040. Proc Natl Acad Sci U S A. 1999. PMID: 10097159 Free PMC article.
-
Functional cloning and reconstitution of vertebrate odorant receptors.Life Sci. 2001 Apr 6;68(19-20):2199-206. doi: 10.1016/s0024-3205(01)01006-2. Life Sci. 2001. PMID: 11358328
-
Molecular and cellular basis of human olfaction.Chem Biodivers. 2004 Dec;1(12):1857-69. doi: 10.1002/cbdv.200490142. Chem Biodivers. 2004. PMID: 17191824 Review.
Cited by
-
Eugenol inhibits the GABAA current in trigeminal ganglion neurons.PLoS One. 2015 Jan 30;10(1):e0117316. doi: 10.1371/journal.pone.0117316. eCollection 2015. PLoS One. 2015. PMID: 25635877 Free PMC article.
-
How does your kidney smell? Emerging roles for olfactory receptors in renal function.Pediatr Nephrol. 2016 May;31(5):715-23. doi: 10.1007/s00467-015-3181-8. Epub 2015 Aug 12. Pediatr Nephrol. 2016. PMID: 26264790 Free PMC article. Review.
-
Heterodimerization and surface localization of G protein coupled receptors.Biochem Pharmacol. 2007 Apr 15;73(8):1043-50. doi: 10.1016/j.bcp.2006.09.001. Epub 2006 Sep 9. Biochem Pharmacol. 2007. PMID: 17011524 Free PMC article.
-
Odor maps in the olfactory cortex.Proc Natl Acad Sci U S A. 2005 May 24;102(21):7724-9. doi: 10.1073/pnas.0503027102. Epub 2005 May 23. Proc Natl Acad Sci U S A. 2005. Retraction in: Proc Natl Acad Sci U S A. 2010 Oct 5;107(40):17451. doi: 10.1073/pnas.1000897107. PMID: 15911779 Free PMC article. Retracted.
-
Vapor detection and discrimination with a panel of odorant receptors.Nat Commun. 2018 Nov 1;9(1):4556. doi: 10.1038/s41467-018-06806-w. Nat Commun. 2018. PMID: 30385742 Free PMC article.
References
-
- Araneda RC, Kini D, Firestein S. The molecular receptive range of an odorant receptor. Nat Neurosci. 2000;3:1248–1255. - PubMed
-
- Belluscio L, Gold GH, Nemes A, Axel R. Mice deficient in Golf are anosmic. Neuron. 1998;20:69–81. - PubMed
-
- Breer H, Boekhoff I. Second messenger signalling in olfaction. Curr Opin Neurobiol. 1992;2:439–443. - PubMed
-
- Brunet LJ, Gold GH, Ngai J. General anosmia caused by a targeted disruption of the mouse olfactory cyclic nucleotide-gated cation channel. Neuron. 1996;17:681–693. - PubMed
-
- Buck LB. Information coding in the vertebrate olfactory system. Annu Rev Neurosci. 1996;19:517–544. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous