

Adenosine A1 receptors reduce release from excitatory but not inhibitory synaptic inputs onto lateral horn neurons
- PMID: 11487654
- PMCID: PMC6763129
- DOI: 10.1523/JNEUROSCI.21-16-06308.2001
Adenosine A1 receptors reduce release from excitatory but not inhibitory synaptic inputs onto lateral horn neurons
Abstract
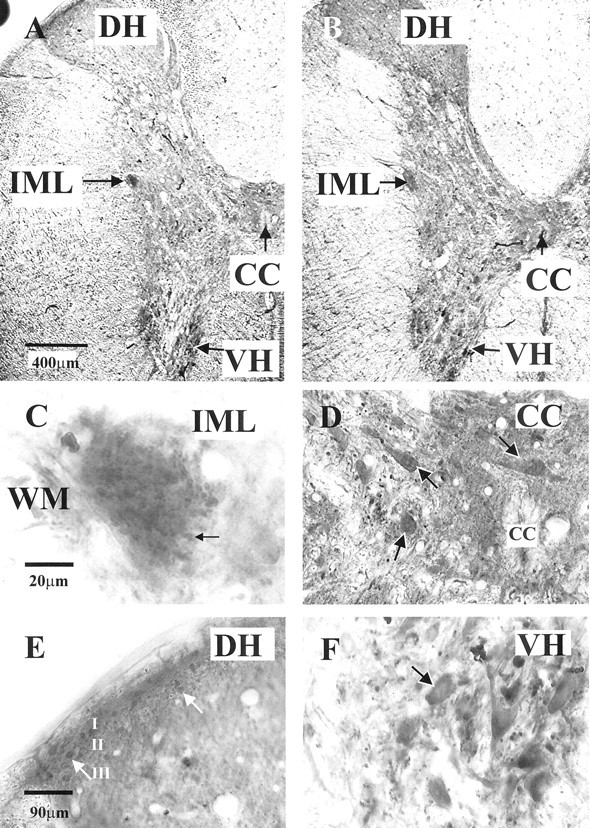
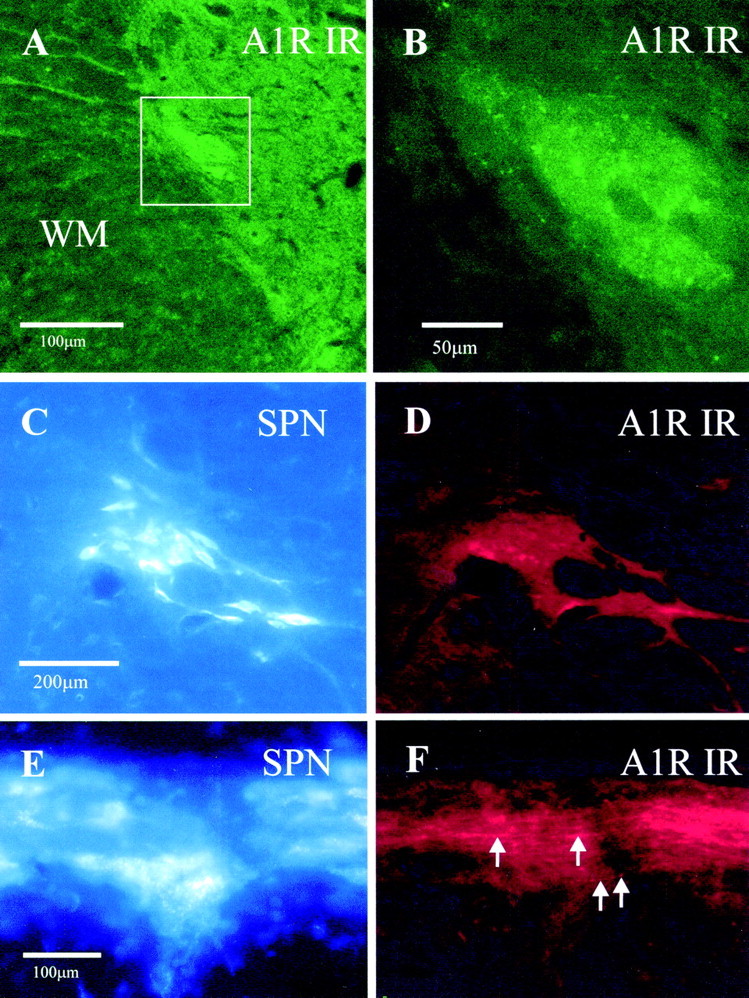
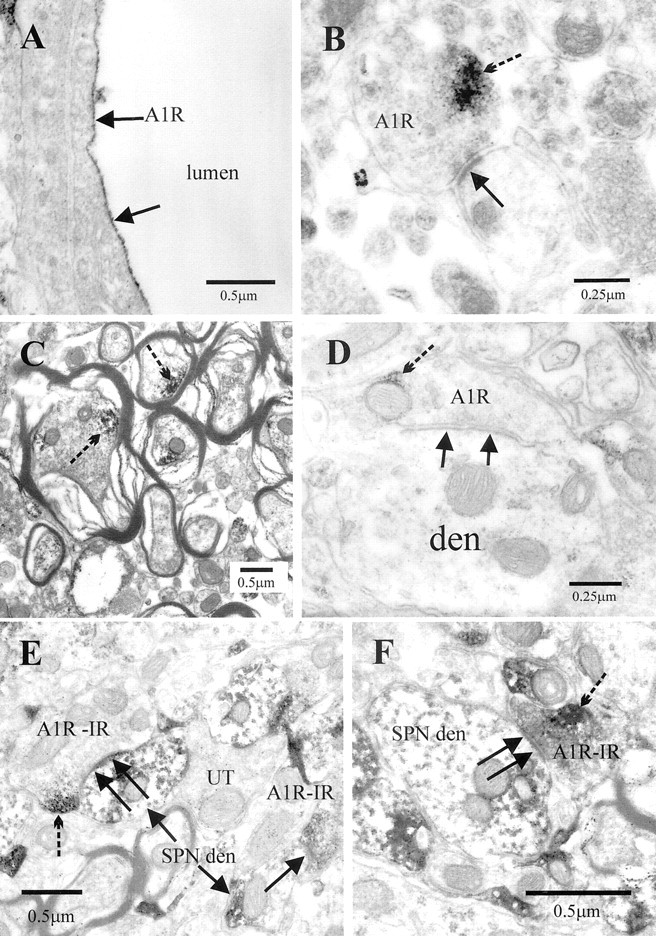
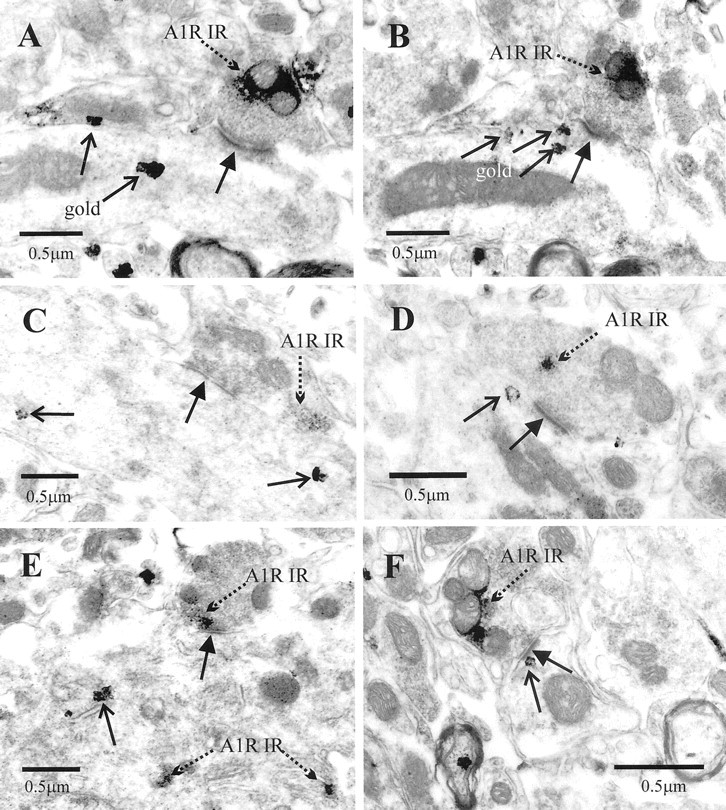
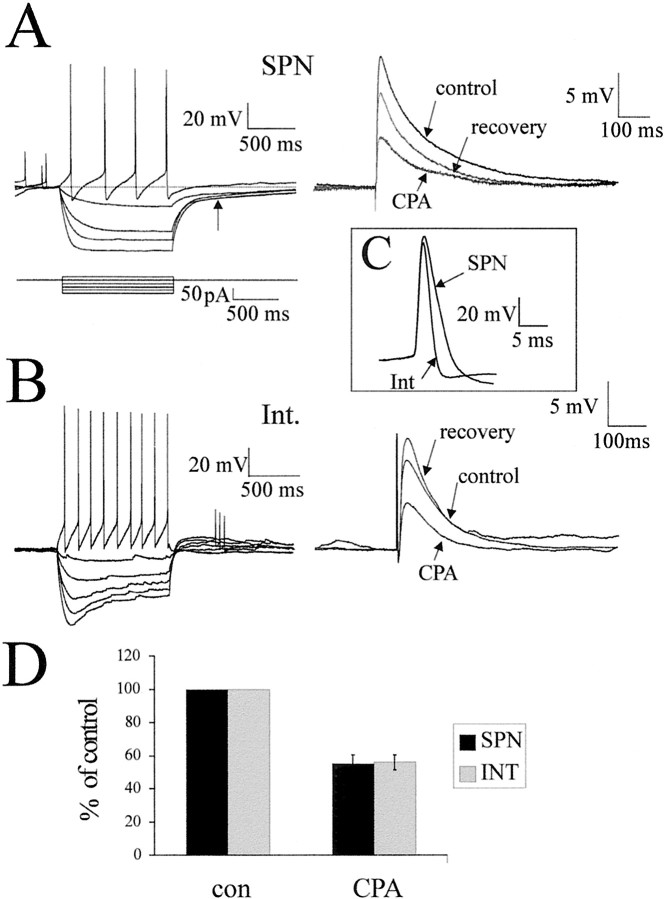
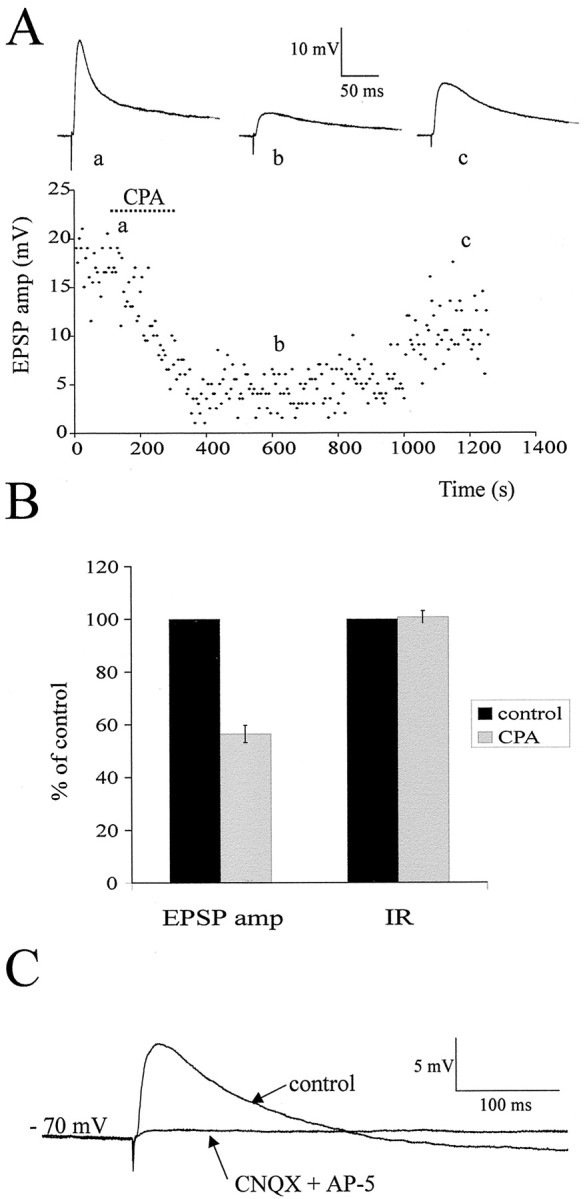
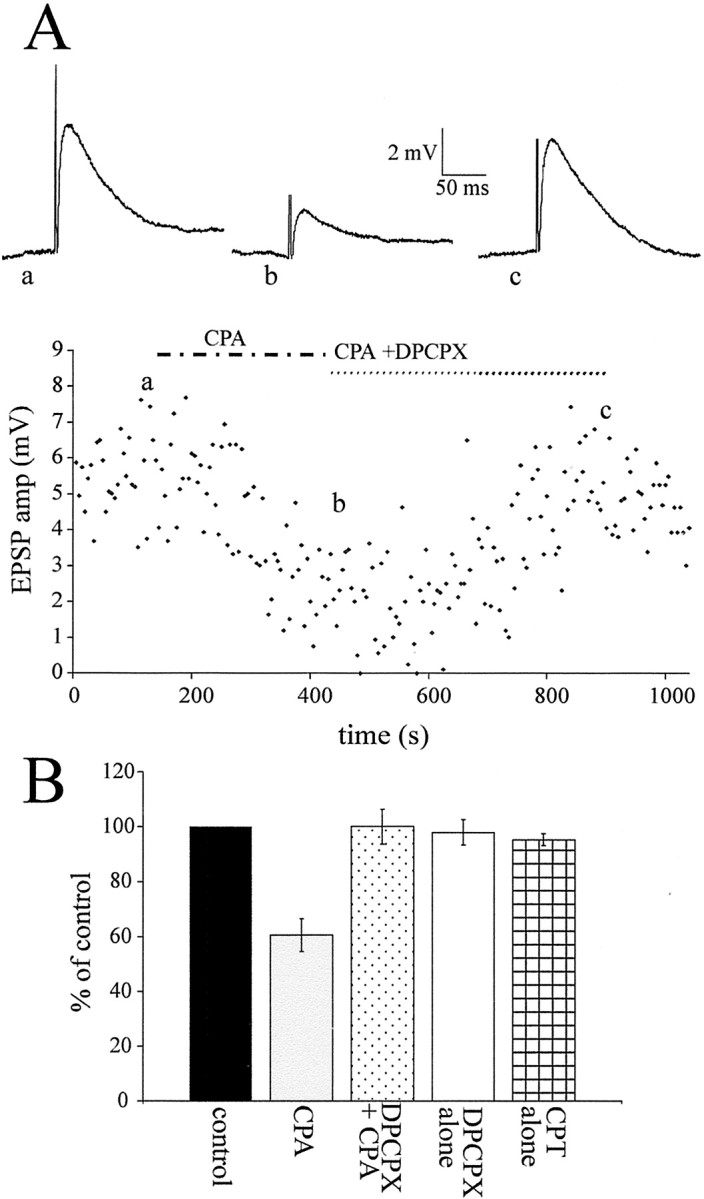
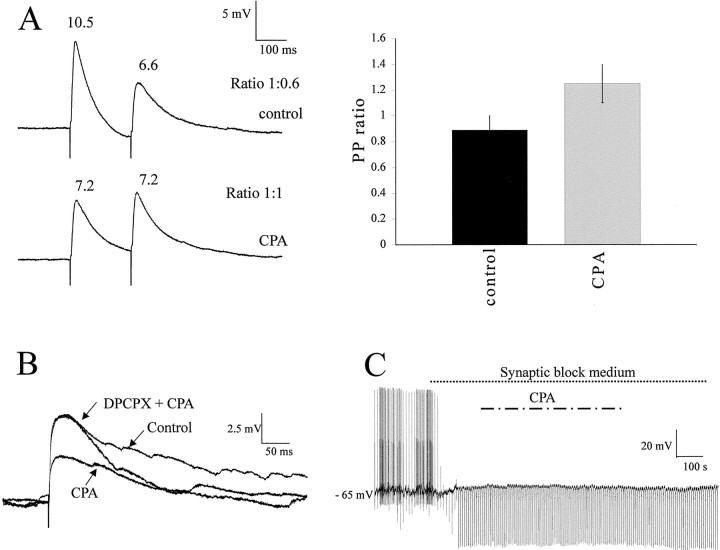
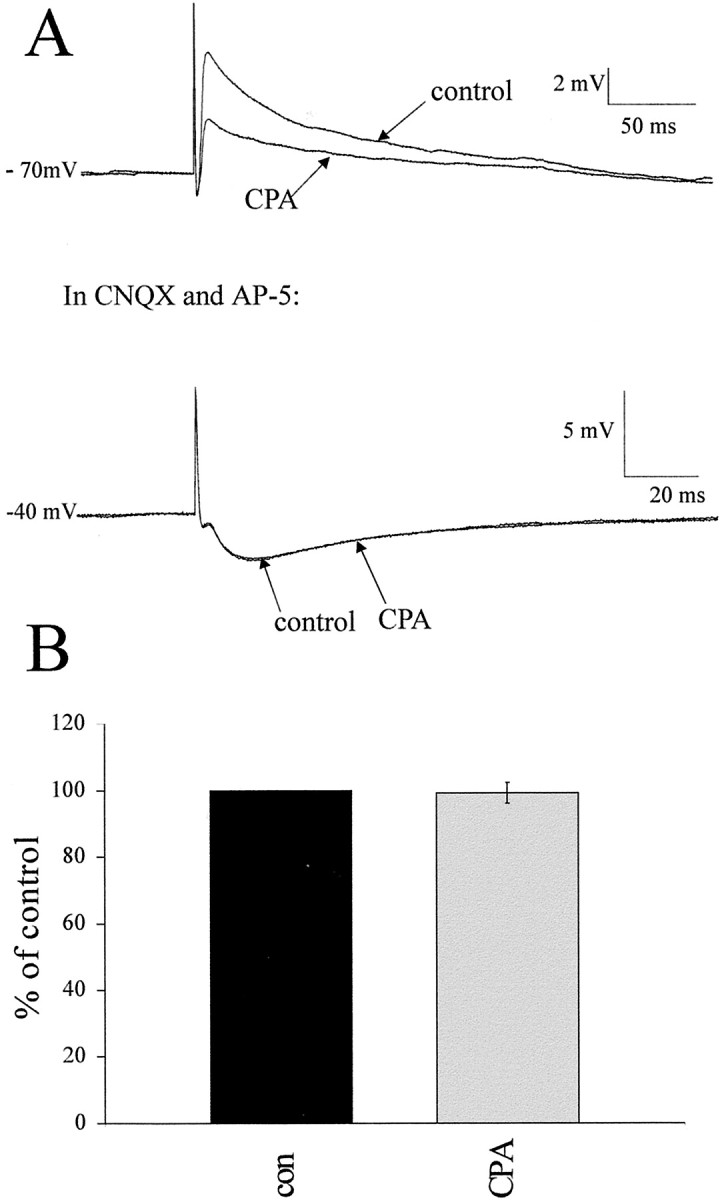
Although adenosine is an important neuromodulator in the CNS, its role in modulating sympathetic outflow at the level of the spinal cord has not been studied. Because very little is known about adenosine A1 receptors (A1Rs) in the spinal cord, we determined their location and role with particular reference to the control of sympathetic preganglionic activity and interneuronal activity in the rat. High levels of immunoreactivity for A1Rs were observed throughout the spinal cord. Immunostaining was dense in the intermediolateral cell column (IML) and intercalated nucleus, regions containing retrogradely labeled sympathetic preganglionic neurons (SPNs). Electron microscopy revealed A1R immunoreactivity (A1R-IR) within presynaptic terminals and (to a lesser extent) postsynaptic structures in the IML, as well as the luminal membrane of endothelial cells lining capillaries. Using double-labeling techniques, some presynaptic terminals were observed to synapse onto SPNs. To investigate the effects of activating these A1Rs, visualized whole-cell patch-clamp recordings were made from electrophysiologically and morphologically identified SPNs and interneurons. Applications of the A1R agonist cyclopentyladenosine (CPA) reduced the amplitude of EPSPs elicited by stimulation of the lateral funiculus, an effect blocked by the A1R antagonist 8-cyclopentyl-1,3-dipropylxanthine. These effects were attributable to adenosine acting at a presynaptic site because CPA application increased the paired-pulse ratio. CPA did not affect evoked IPSPs. These data show that activating A1Rs reduces fast excitatory, but not inhibitory, transmission onto SPNs and interneurons in the IML and that A1Rs may play a protective role on neurons involved in the control of sympathetic outflow.
Figures
References
-
- Ackley MA, Baldwin SA, King AE. Inhibition of adenosine uptake attenuates glutamatergic synaptic transmission in lamina II of the rat spinal cord in vitro. Soc Neurosci Abstr. 2000;26:821.5.
-
- Alkadhi KA, Brown TR, Sabouni MH. Inhibitory effect of adenosine on transmission in sympathetic ganglia. Naunyn Schmiedebergs Arch Pharmacol. 1984;328:16–19. - PubMed
-
- Chau D, Johns DG, Schramm LP. Ongoing and stimulus-evoked activity of sympathetically correlated neurons in the intermediate zone and dorsal horn of acutely spinalized rats. J Neurophysiol. 2000;83:2699–2707. - PubMed
-
- Choca JI, Proudfit HK, Green RD. Identification of A1 and A2 adenosine receptors in the rat spinal cord. J Pharmacol Exp Ther. 1987;242:905–910. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources